
ХАРАКТЕРИСТИКА РАЙОНА и ОБЪЕКТА ИССЛЕДОВАНИЙ
Рис. 1 Устье безымянной реки, впадающей в залив Ахматова с западного берега. На переднем плане дельта реки, на правом склоне долины – 20-метровая, на левом 40-50 – метровая морские террасы. Рис. 2 Долина безымянной реки, впадающей в залив Ахматова с западного берега. Вид сверху – вниз. В середине подмываемое рекой обнажение чёрных – серых глин, увеличивающих видимую мощность с 4 до 7 м вверх по течению реки. Виден неровный контакт глин и налегающих на них гравийно-песчано-галечных отложений. 

ФАКТИЧЕСКИЙ МАТЕРИАЛ
09 августа 2020 г. Западный берег залива Ахматова, устье ручья в виде лопастной дельты выдвижения. Долина ручья – реки уходит в горы. (79°04’04,6” с.ш 102°44’43,0” в.д.). Здесь примерно в 400 м от края дельты река подмыла правый берег и обнажила разрез алевритов с вертикальной стенкой. Высота стенки до 8 м. Обнажение сложено горизонтально-слоистыми серо-жёлтыми алевритами. Слоистость обеспечена цветом – переслаиваются жёлтые и серые слои. Серые до 2 см, желтые до 5 см. Разрез не расчищен полностью из-а нехватки времени. Поэтому произведено беглое описание обнажения. На высоте 1,5–1.8 м от подошвы обнажения (дна долины) в алевритах залегает заметный прослой мелких галек толщиной 25 см. Отличается тем, что галька в нём уложена хаотично. На высоте 6 м произведена расчистка для отбора створок раковин моллюсков и породы для ЭПР-анализа. Здесь также есть прослой мелкой гальки толщиной около 2 см, кроме того, есть отдельные гальки в породе. Встречаются тонкостенные раковины, которые невозможно отобрать без разрушения. Большинство из них имеет очень переливчатый и разноцветный перламутровый слой. Раковины двустворчатые. Произведён отбор образца этих раковин и осколков более толстостенных раковин с породой – образец 3298/1. Выше слоя отбора в 0,3 м (на высоте 6,5 м) в алевритах появляются прослои растительности чёрного цвета и эти прослойки в первые миллиметры-сантиметры встречаются и выше – до вершины стенки. Это тонкие спресованные, видимо волокна водорослей. Отобран образец растительности с породой 3298/2. Произведён осмотр отложения выше по течению ручья и обнаружен слой сильно пахнущей (сероводородом?) чёрной глины, которая и красит воду в ручье в тёмно-серый цвет. Там же есть продолжение разреза вверх. На дне долины найдены 2 влажные лепёшки тех самых чёрных водорослей, которые мелкими залежами отобраны в обр. 3298/2. Лепёшки до 2 см толщиной, и они практически состоят из органики. Образец 3298/4 – чёрные водоросли из верхнего отрезка долины. Окатыши породы с водорослями находятся на днище долины. При их разламывании чувствуется запах йода. На дне долины найдены многочисленные створки раковин: Macoma, Hiatella, и ещё какая-то удлинённая раковина. Они отобраны в образец 3298/3. На дне долины также многочисленные кости: 2 позвонка кита, несколько рёбер, по-видимому моржа средней сохранности. Они окатаны при перемещении из разреза и имеют типичный белый цвет. Найден также один клык моржа отличной сохранности, с эмалью, коричневатого цвета с частью челюсти. 13 августа 2020 г. Продолжение описания обнажения на западном берегу залива Ахматова (79°04’11,3” с.ш 102°43’33,0” в.д.) точка находится в глубине долины на расстоянии 890 м от края дельты реки (берега моря). Высота подошвы обнажения по эклиметрированию – 25 м, высота по барометру 25 м, по GPS - 23 м. Подножие обнажения I, описанного 9 августа, расположено на высоте 9 м. Обнажение II с подошвой на высоте 25 м характерно отличной вскрытостью ручьём стенки чёрных глин, которые перекрываются песчано-галечно-гравийной толщей. Выше по течению глины увеличивают видимую мощность с 4 до 7 м и далее выклиниваются вверх по долине, закрываясь осыпными шлейфами песчано-галечной толщи, слагающей террасу высотой 51 м в бровке, 60 м в тыловом шве. На поверхности террасы залегают хорошо и отлично окатанные гальки. Чёрные глины вскрыты ручьём под левым склоном долины и образуют выпуклую стенку высотой 3,5–4 м. Они представлены алевритовой глиной чёрного цвета, которая, быстро окисляясь меняет цвет на серый. В этой стенке происходит активное обваливание обнажения пластинками в результате нагревания, особенно под лучами солнца. Это происходит потому, что глины насыщены льдом (до 20–30%), причём ледяными вертикальными жилками в виде многозубцев, обращённых остриями вверх и с общим льдистым основанием. Высота такого отдельного тела 0,7 м, ширина 0,6 м. Алевритовые глины отлично слоисты, толщина слойков от первых миллиметров до 1 см. Но в них есть некоторая неоднородность, что придаёт глинам воздушность при сминании. Вероятнее всего это происходит из-за наличия в глинах остатков растительности, а именно волокон водорослей, отпечатки и сами тонкие волокна которых иногда обнаруживаются при разделении слойков. Ручей на урезе воды и выше на 0,5 м вскрыл ракушняковую банку, постоянно препарируя и вымывая раковины двустворчатых моллюсков. Раковины расположены в беспорядочном положении, которые вертикально, которые горизонтально или наклонно залегают по отношению к слоистости. Или сами слои имеют нарушение горизонтального залегания, чего в этой части обнажения не определить. Часто раковины прислонены друг к другу. Они заполнены льдистой породой или льдом, отчего кажущиеся прочными, быстро разрушаются при вытаивании льда и с потерей контакта с находящейся внутри створок породой. Есть раковины полные из двух створок. Несомненно, мерзлотные процессы ответственны за трещиноватость и разрушение раковин. Раковины двустворчатых моллюсков в глинах тонкостенные, в отличие от толстостенных, повсюду раскиданных по дну долины. Из этой части обнажения в интервале от уреза воды до 0,5 м выше него отобраны образцы: 3298/5 – порода -алевритовая глина со створками раковин моллюсков для ЭПР-анализа возраста в п-эт. пакет. 3298/6 – глины для микрофаунистического и диатомового анализа в п-эт. пакет, 3298/7 – раковины для определения видов моллюсков в 2 п-эт. пакета. Последняя проба собиралась так, чтобы в раковинах оставалась скрепляющая их глина. Но в пакете произошло вытаивание льда и раковины разрушились. Судя по этим пробам влажность глин высока – не менее 20%. Все пробы превратились в тестоподобную полужидкую массу. Но во время отмывки раковин уже в ноябре 2020 г. удалось получить 3 полные двустворки с заполняющим их глинистым алевритом, несколько одиночных створок и осколки разного размера. Вся проба отдана палеонтологу А. В. Крылову для определения видового состава двустворчатых моллюсков. Из нескольких одиночных створок удалось изъять заполняющий их глинистый алеврит в отдельный пакет 3298/7 для других видов анализа. Выше по долине на 50-100 м и при высоте уреза воды 26,7 м над уровнем моря глины вскрываются на большую мощность – до 7 м. Из этой стенки уже серых окислившихся глин отобраны пробы с горизонта 29,2 м над у.м. (2,5 м над урезом воды в ручье) – образец 3298/8, и с высоты 33,7 м, из кровли глин – обр. 3298/9. Кровля глин неровная, но без всяких затёков и карманов. Выше залегает толща переслаивания гравия и пластов серого песка, желтовато-серого алеврита и растительности (вероятно, водорослей чёрного цвета. Эти пачки состоят из растительности толщиной прослоев до 2–3 см, слойков алевритов до 10 см и песков с гравием толщиной в первые сантиметры. Пачка в целом имеет толщину 15-20 см. Таких пачек над глинами среди гравийников насчитывается 3, в некоторых местах на склоне – 4. Так как осыпной склон имеет крутизну 30-35° и сложен галечно-гравийным материалом все слои осадков смещены вниз по склону, а эти пачки перереслаивания водорослей, песков и алевритов без разрыва сплошности, но вскрываются они слегка на разной высоте, изменяя её в пределах 1-2 м. Контакт гравийников с подстилающими глинами неровный. Гравийники постоянно надвигаются на бровку стенки, сложенную глинами, и осыпаются далее до дна ручья. Из зоны контакта, на 0,8 м выше глин (34.5 м над ур. моря) из гравийников взят образец 3298/10. На высоте 2 м выше контакта с глинами (35,7 м над ур. моря) отобраны образцы из пачки переслаивания песков, алевритов, водорослей: 3298/11 – алеврит с гравием в пакет, 3298/11 ОСЛ- алеврит с гравием в трубку для проведения ОСЛ-анализа возраста, 3298/12 древесины из пачки переслаивания для радиоуглеродного датирования Образец 3298/13 – кость морского млекопитающего (ребро?) из осыпи на этом же высотном уровне. Древесный ствол диаметром 6 см торчал из пачки переслаивания водорослей, алевритов, песков на 1,2 м. Произведено углубление до мерзлоты в стенке (на, 0,4 м) и извлечена погребённая часть ствола. Древесина старая и отслаивается по годичным кольцам как кора. Последующая распиловка древесины показала, что древесина находится в стадии разложения (пилится как слегка трухлявое дерево), но сохраняет структуру годичных колец, особенно в извлечённой погребённой и оттаявшей части. В данном месте на склоне насчитывается ещё две таких пачки переслаивания, съезжающих вниз по осыпному склону. До бровки на высоте 51,7 м над уровнем моря на осыпном склоне встречаются кости морских млекопитающих, которые попадают на дно долины именно из этих гравийников и галечников. Возвращение к обнажению I, из которого 9 августа были отобраны алевриты для ЭПР анализа. Подошва этого обнажения, вскрытого ручьём по правому склону долины, лежит на высоте 9,5 м над уровнем моря. Толща алевритов имеет видимую мощность 10 м. В обращённой к ручью стенке угол склона практически вертикальный. К разрезу можно было подобраться по долинке маленького овражка, отходящего от ручья к западу. Вся толща алевритов более или менее однородная представлена переслаиванием серых и желтовато-серых слоёв алевритов. Иногда в толще содержатся прослойки-линзы галечников толщиной до 1–2 см (мелкая галька и гравий). Образец 3298/14 алевритов отобран с глубины 1 м ниже бровки обнажения (18, 5 м над у.м.) В 1 м ниже бровки в алевритах попадаются целые раковины моллюсков и их осколки, очень тонкие, разваливающиеся при извлечении из осадка. Есть раковины двустворчатых моллюсков и гастропод. В горизонте 18, 5 м над уровнем моря собраны тонкие раковины двустворчатых моллюсков в пробирку – образец 3298/15. На невскрытой поверхности обнажения есть белые и серые выцветы солей по вкусу поваренной соли – обр. 3298/16 с горизонта 18,5 м. На высоте 17,5 м найдена раковина гастроподы диаметром 1 см, хрупкая – отобрана в пластиковую пробирку – обр. 3298/17. Это горизонт, из которого 9 августа был отобран образец 3298/1. Отсюда же отобран образец алевритов 3298/18 в трубку для проведения ОСЛ-анализа возраста. С высоты 16 м отобраны соли, которые прослеживаются в разрезе как прослой толщиной до 1 см. Соли представлены прозрачными кристаллами с солёным и противным вкусом. Из них взят образец 3298/19. Нижняя часть обнажения рассмотрена со стороны вертикальной эрозионной стенки ручья. Она характерна переслаиванием серых алевритов большей частью (слойки по 3–10 см) и прослоями алевритом рыжего цвета по 1–2 см толщиной. На высоте 2 м над подошвой обнажения (11,5 м над у.м.) залегает прослой мелкого галечника толщиной 15–20 см. В подошве галечника есть прослойки гравия и песка, из которых взят образец 3298/20. Гальки расположены прослоями, падающими в разные стороны под большими углами. Из основания уступа размыва с высоты 0,4 м (9,9 м над у.м.) взят образец алеврита 3298/21. В стенке на высоте примерно 5 и 6,5 м над основанием (14,5 и 16 м над у.м.) видны тонкие прослойки солей, которые отобраны из стенки в распадке (обр. 3298/19). В целом разрезы показали, что морские отложения состоят из 3 пачек осадков. По высоте самая низкая пачка отложений – алевритовая, располагается ближе к морю (9,5–20 м). Средняя – чёрные глины на высоте 25–33,7 м, высокая – гравийно-галечная толща (33.7–50 м). Высоты определены эклиметрированием и по данным прибора GPS. Значит их точность лежит в пределах 1–2 м. Рис. 1 Схема обнажения 3298, изученного в долине реки, впадающей в залив Ахматова с западного склона. Условные обозначения: 1 – алеврит, 2 – галька, 3 – прослой соли, 4 – песок, 5 – гравий, 6 – глина, 7 –раковина моллюска, 8 – граница стратиграфического несогласия, 9- плавник древесины, 10 – кость млекопитающего, 11 – номер образца Рис. 2 Разрез (I) морских алевритов в 400 м от края дельты безымянной реки, впадающей в залив Ахматова Рис. 3 Хорошо окатанная галька на поверхности морской террасы высотой 50–60 м на западном склоне залива Ахматова Рис. 4 Раковины двустворчатых моллюсков в чёрных глинах в основании разреза II Рис. 5 Размыв ручьём чёрных глин с банкой двустворчатых моллюсков (разрез II) 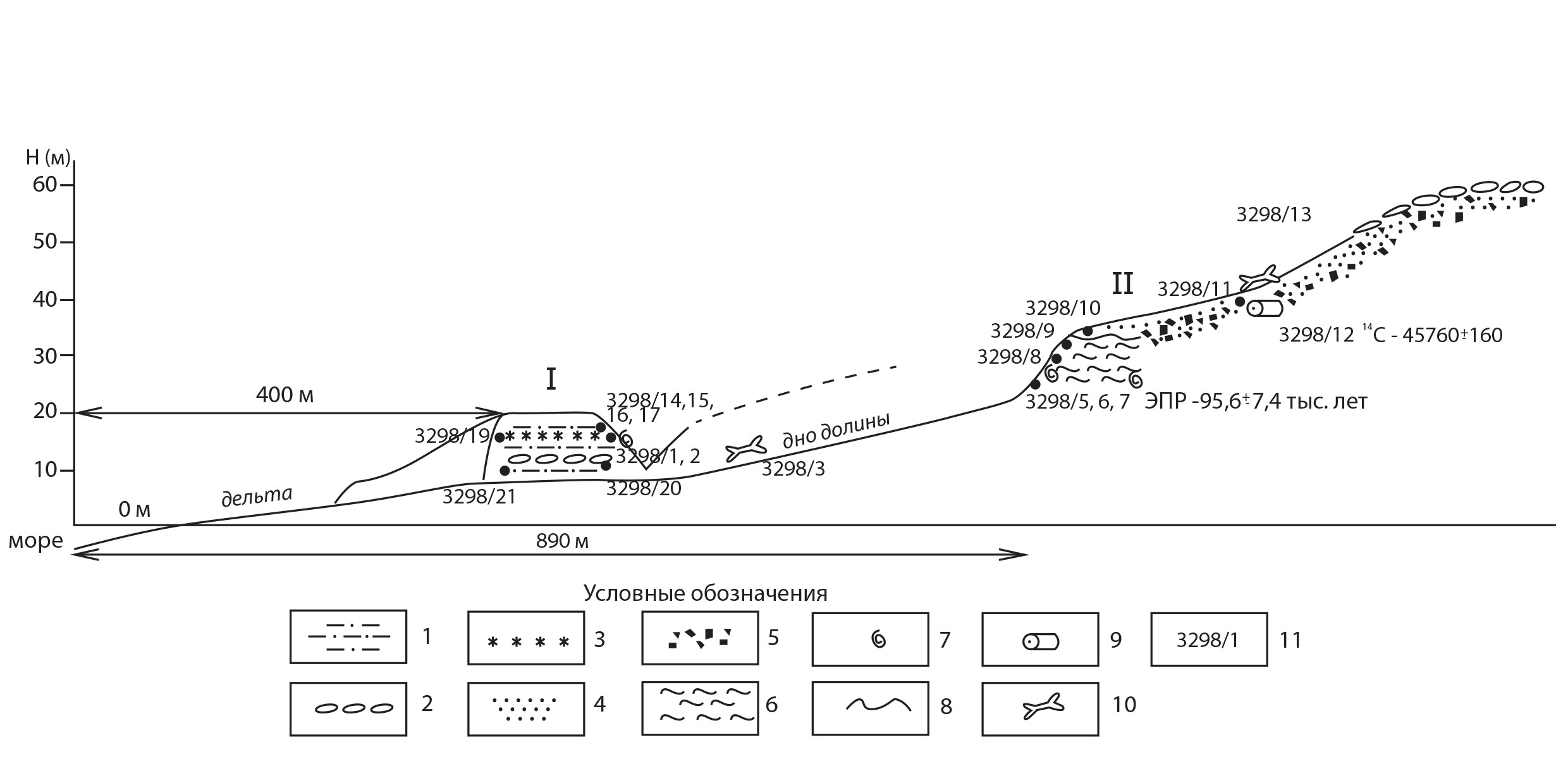




ДИАТОМОВЫЙ АНАЛИЗ
Господствующую группу диатомей в изученных отложениях составляют морские диатомовые водоросли, число которых составляет 28 видов (Actinocyclus sp., Chaetoceros sp., Diploneis subcincta, Thalassiosira gravida, Trachyneis aspera и другие). В диатомовых комплексах часто встречены солоноватоводно-морские виды Cocconeis scutellum, Diploneis smithii, Grammatophora angulosa и другие, солоноватоводные представлены «единично» Bacillaria socialis, Thalassiosira baltica, также обнаружены пресноводные Fragilaria spp., Pinnularia obscura, Ulnaria ulna и другие. Большинство установленных диатомовых водорослей являются космополитами и практически все известны в современных диатомовых биоценозах и в диатомовых комплексах из голоценовых отложений морей Восточной Арктики (Полякова Е. И., 1997; Cremer H., 1999; Цой И. Б., Обрезкова М. С., 2017). В нижней алевритовой пачке отобраны образцы 3298/21, 3298/20, 3298/2, 3298/4, 3298/14, 3298/16, 3298/19. В самом нижнем слое алевритов образец 3298/21 установлены диатомовые комплексы in situ, состоящие из немногочисленных створок морских диатомовых водорослей, также присутствуют цисты золотистых водорослей. Диатомеи представлены как относительно грубоокремненными створками морских диатомовых, так и тонкоокремененными створками, маленького размера (Fragilaria spp.; длина около 15 микрон) хорошей сохранности. Вероятно, эти тонкоокремненные створки пресноводных диатомей были занесены в отложения позднее, поскольку створки морских диатомей встречены и в обломочном состоянии и мало, вероятно, чтобы такие «тонкие» створки пресноводных могли сохраниться целыми. В образце 3298/21 наиболее часто встречены диатомовые водоросли: неритические планктонные Aсtinocyclus sp., Bacterosira fragilis, Chaetoceros sp., Thalassiosira gravida, океаническая Rhizosolenia styliformis; ледово-морские планктонные Fragilariopsis oceanica; бентосные Amphora sp., Cocconeis costata, C. scutellum, Grammatophora angulosa, Diploneis smithii, Rhabdonema sp., Trachyneis aspera и другие. Присутствие хотя и немногочисленных створок планктонных диатомовых позволяет сделать вывод о формировании отложений, вмещающих этот диатомовый комплекс, в прибрежных относительно глубоких условиях морского бассейна. В образце 3298/20 из песчано-галечных отложений установлены единичные створки морских планктонных диатомей Thalassiosira gravida, и обломки створок морских и бентосных диатомей, единично обнаружена створка Rhabdonema cf. japonicum, возможно, переотложенная из неогеновых отложений. Выше по разрезу этой пачки алевритов: в образцах 3298/2, 3298/14, 3298/16 и 3298/19 установлены обедненные количественно морские диатомовые комплексы, в которых наиболее заметны: бентосные «редко» Grammatophora angulosa, Diploneis smithii, D. subcincta, Cocconeis costata, C. scutellum; планктонные «единично» Chaetoceros sp., Thalassiosira gravida, T. nordenskioеldii, Porosira glacialis и другие. Установленные диатомовые водоросли позволяют сделать вывод о формировании отложений в сублиторальной зоне морского бассейна, солоноватоводные и солоновато-морские диатомеи характеризуют некоторое опреснение этой сублиторальной зоны. Таким образом, в обнажении I, в нижней части алевритовой пачки установлен морской диатомовый комплекс, определяющий прибрежные условия, возможно, более глубоководные, поскольку планктонная группа диатомей в диатомовых комплексах заметнее и по видовому составу и количественно. Выше по разрезу диатомовые комплексы очень похожи по видовому составу, но бентосные виды более разнообразны в видовом отношении и более представительны количественно и характеризуют морские сублиторальные условия. Выше по разрезу в пачке глин образцах 3298/6, 3298/8, 3298/9 и 3298/10 установлены морские обедненные в видовом и количественном отношении диатомовые комплексы, в образце 3298/9 обнаружено наиболее количество створок диатомей и количество видов. В диатомовых комплексах встречены: неритические планктонные Thalassiosira gravida, ледово-морские планктонные Fragilariopsis cylindrus, Melosira arctica, бентосные Cocconeis costata, C. scutellum, Diploneis smithii, Grammatophora angulosa, Navicula directa, Pseudogomphonema cf. septentrionale и другие, обломки створок бентосного вида Trachyneis aspera. Можно предположить, что диатомовые створки накапливались в сублиторальной зоне морского бассейна. В образце 3298/8 обнаружен диатомовый комплекс, состоящий из морских диатомей, представленный единичными целыми створками, в большинстве же диатомеи представлены обломками, которые возможно идентифицировать, и очень маленькими фрагментами створок. Можно предположить, что диатомовые створки накапливались в сублиторальной зоне морского бассейна, возможно, в его прибойной части, чем предположительно можно объяснить их плохую сохранность. В образце 3298/11 установлены только обломки створок морских диатомей. При сравнении диатомовых водорослей из пачек (пачка I) алевритов и глин (пачка II) можно заключить, что верхняя часть алевритов и глины формировались в сублиторальной части морского бассейна, и только нижняя часть алевритов (образец 3298/21), вероятно, формировалась в более глубоководных условиях. Таблица 1. Диатомеи в разрезе 3298 архипелаг Северная Земля, остров Большевик Диатомовые водоросли № образцов 3298-21 3298-20 3298-19 3298-2 3298-14 3298-16 3298-4 3298-6 3298-8 3298-9 3298-10 3298-11 Океанические и неритические Aсtinocyclus sp. vr vr vr Bacterosira fragilis Gran vr fr. Chaetoceros septentrionalis Østrup vr Ch. sp. vr vr vr vr Fragilariopsis cylindrus (Grunow) Krieger vr F. oceanica (Cleve) Hasle vr Hyalodiscus obsoletus Sheshukova vr Melosira arctica (Ehrenberg) Dickie vr Porosira glacialis (Grunow) Jorgensen vr Rhizosolenia styliformis Brighwell vr Thalassiosira baltica (Grunow) Ostenfeld T. decipiens (Grunow) Jorgensen vr T. gravida Cleve f vr vr vr vr vr vr vr vr vr vr T. nordenskioеldii Cleve vr Centricae fr. fr. fr. fr. fr. fr. fr. fr. fr. fr. fr. fr. Бентические морские Amphora proteus Gregory fr. fr. Bacillaria socialis (Gregory) Grunow fr. fr. fr. fr. Cocconeis costata Gregory fr. fr. C. scutellum Ehrenberg r vr vr r vr vr vr vr vr Diploneis interrupta (Kützing) Cleve vr vr D. smithii (Brébisson) Cleve vr vr vr vr fr. vr vr vr vr fr. D. subcincta (Schmidt) Cleve vr vr fr. fr. vr fr. Fallacia pygmaea (Kützing) Stickle & D.G. Mann vr Grammatophora angulosa Ehrenberg vr f vr vr vr r vr Gyrosigma sp. fr. Hannaea arcus (Ehrenberg) Patrick fr. Диатомовые водоросли № образцов 3298-21 3298-20 3298-19 3298-2 3298-14 3298-16 3298-4 3298-6 3298-8 3298-9 3298-10 3298-11 Lyrella lyra (Ehrenberg) Karaeva vr vr vr Navicula directa (Smith) Ralfs vr vr vr N. giacialis (Cleve) Grunow fr. N. transistans Cleve vr N. sp. vr vr vr vr fr. Nitzschia sp. vr fr. fr. fr. fr. vr Pleurosigma sp. fr. Pseudogomphonema cf. septentrionale (Østrup) Medlin vr vr Rhabdonema sp. (септы) vr vr fr. Trachyneis aspera (Ehrenberg) Cleve vr fr. fr. fr. fr. fr. fr. vr fr. Pennatae fr. fr. fr. fr. fr. fr. fr. fr. fr. fr. fr. fr. Бентические пресноводные Amphora sp. vr Fragilaria spp. r r vr Pinnularia obscura Krasske vr Psammothidium sp. vr Ulnaria ulna (Nitzsch) Compère fr. fr. fr. fr. Третичные диатомеи Rhabdonema cf. japonicum Tempère & Brun vr Chrysophycean cysts vr vr vr vr vr Примечание. vr - very rare - единично (1-5 створок на препарат) r - rare - редко (6-30) f – frequently - нередко (31-60); fr. – обломки створок диатомей
МИКРОФАУНА
В образцах №№ 3298/1 и 3298/2 определены в умеренном количестве агглютинирующие фораминиферы Saccammina sp. Этот вид имеет широкое возрастное распространение от силура до ныне. Эти фораминиферы «… почти повсеместно распространены в мезозое и кайнозое, расселены в современных морях, образуя в холодноводных условиях массовые скопления». [Введение в изучение фораминифер. Под редакцией Н. Н. Субботиной, Н. А. Волошиновой, А.Я. Азбель, 1981г.]. Образец №3298/6 содержит достаточно многочисленный аркто-бореальный комплекс, в котором 26 видов и 796 экземпляров раковинок фораминифер в основном хорошей и средней сохранности. В этом образце явное преобладание кассидулинид, среди которых преобладает вид Cassandra teretis с хорошо развитыми крупными раковинками. Однако попадаются среди них мелкие, деформированные и уродливые (сросшиеся) особи. Этот арктический вид обитает в пределах материковой отмели. Меньше раковинок Cassidulina reniforme и C. subacuta. Ретроэльфидиумы малочисленны и представлены мелкими, часто ломаными раковинками одного вида Retroelphidium atlanticum. Нониониды Cribrononion obscurus, Cr.incertus, Nonionelina labradorica и Melonis zaandamae единичны, только вид Astrononion gallowayi достаточно многочисленен (74 экземпляра) и имеет мелкие, в основном ломаные раковинки. Выделены также хайнезины вида Haynesina orbiculare. Из криброэльфидиумов определены Cribroelphidium goesi. Присутствуют букцеллы видов Buccella frigida и B. hannai arctica, определены единичные Pseudopolimorphina novangliae, Hyalinea sp., Asterigerinata pulchella, Alabaminoides mitis, Gavelinopsis praegeri, Bolivina pseudopunctata, Fursenkoina gracilis и другие. Единственная Quinqueloculina sp. имеет ломаную, потёртую раковинку и является, вероятней всего, переотложенной. Присутствующий в этом комплексе вид Cassidulina reniforme является мелководно-морским, а его достаточно большое количество свидетельствует о неблагоприятных условиях для жизни других фораминифер. Это подтверждается и мелкими размерами большинства остальных раковинок комплекса, а также присутствием уродливых форм. Исходя из полученных данных, можно предположить, что аркто-бореальный комплекс этого образца формировался в открытом относительно неглубоком морском бассейне в пределах шельфа, c низкой придонной температурой вод и с солёностью близкой к нормально-морской. Образец № 3298/8 содержит немногочисленный аркто-бореальный комплекс, в котором присутствуют 15видов и 58 экземпляров фораминифер в основном хорошей сохранности. В комплексе наблюдается явное преобладание кассидулинид видов Cassidulina reniforme, Cassidulina subacuta, Cassandra teretis. Среди последних преобладают крупные, хорошо сформированные особи. Вид Cassandra teretis распространён в арктической и бореальной областях, но тяготеет к более холодным водным массам в шельфовых морях. Достаточно многочисленны букцеллы видов Buccella frigida и Buccella hannai arctica, меньше Astrononion gallowayi, Alabaminoides mitis. Единично присутствуют Sigmomorphina undulosa, Discorbis deplanatus, Melonis sp. - juv., Haynesina orbiculare, Cribroelphidium subarcticum. Учитывая полученные результаты, можно предположить, что этот несколько обеднённый комплекс формировался, вероятней всего, в не глубоком морском бассейне с нормальной солёностью и отрицательными придонными температурами вод. В образце № 3298/8 обнаружена единственная раковинка остракоды В образцах №№ 3298/9 и 3298/10 присутствуют в небольшом количестве агглютинирующие фораминиферы Rhabdammina sp. и обломки Textularia sp. Эти морские глубоководные виды распространены повсеместно и имеют широкое возрастное распространение от ордовика до ныне. Образцы №№ 3298/14, 3298/20, 3298/21 содержат единичные раковинки фораминифер (от 2 до 4 экземпляров, см. таблицу) в основном хорошей сохранности. Это агглютинирующая Rotaliammina orchaceae и секреционные бентосные Buccella frigida, Retroelphidium atlanticum, Haynesina orbiculare, Cassandra teretis. Эти виды имеют широкое возрастное распространение в кайнозое и можно предположить, что состав и сохранность данного набора фораминифер предполагает нормально-морские условия (арктическое море) для своего обитания.
ФАУНА МОРСКИХ МОЛЛЮСКОВ
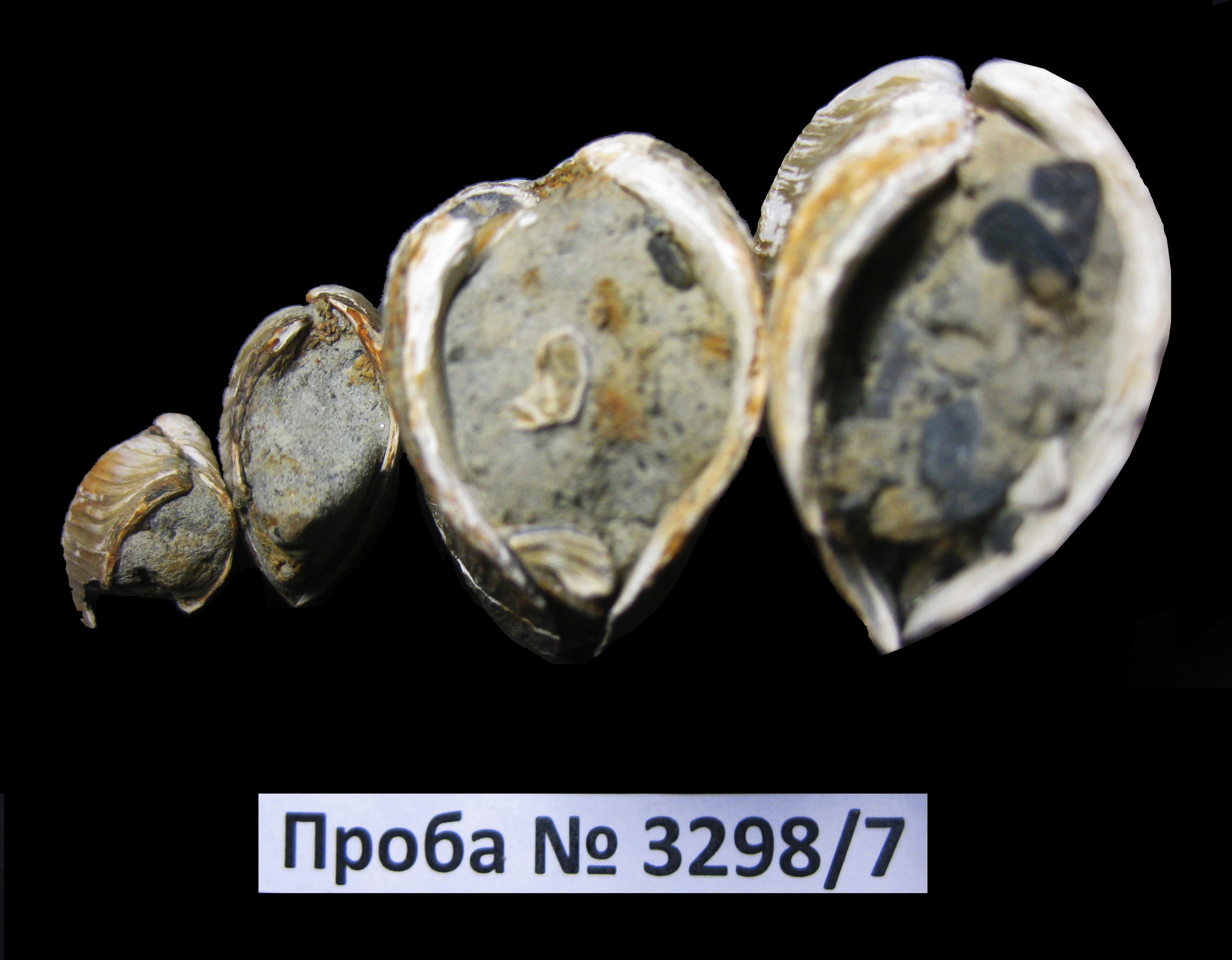
Образец № 3298/7 Mya truncata Linnaeus, 1758 - известен с миоцена, широко распространенный бореально-арктический циркумполярный вид (Петров, 1966, Наумов, 2006, McNeil, 1965). Встречен во всех северных морях России, море Бофорта, у берегов Канадского Арктического архипелага, Гренландии, Исландии, Шпицбергена и Земли Франца-Иосифа. В Атлантическом океане расселяется на юг до Бискайского залива и п-ова Кейп-Код, в Тихом океане – до Желтого моря и залива Пьюджет-Саунд. Стенотермный, умеренно тепловодный, относительно стенобатный, относительно эвритопный, предпочитающий илистые и песчаные грунты вид. Впервые встречается в миоцене Северной Пацифики (McNeil, 1965), встречается в плиоцене Италии и Бельгии (Marquet, 2005). Отмечен на глубинах 5–30 и 80–180 м (реже 0–5 м, 30–80 м и 180–625 м) при температуре от - 2,0 до 13,7°С и солености 23,9 - 35,2‰, в основном на илистых (34% находок) и смешанных (40% находок) грунтах. Миоцен-голоцен (в Арктике ранний плейстоцен-голоцен, преимущественно неоплейстоцен (Мерклин, Зархидзе, Ильина, 1979, Наумов, 2006 и др.) Характеризует мягкие глинистые или песчано-глинистые грунты, глубины арктический-бореальный климат. Абсолютное датирование мий (как и циртодарий и иных зарывающихся моллюсков) не всегда удачно отражает возраст вмещающих отложений из-за химического засорения раковин, однако, в ряде случаев (открытого моря, стабильного режима солености и удаленности от берега и хорошей сохранности самих раковин, захороненных неподалеку от места своего обитания) может быть успешным. Hiatella arctica (Linnaeus, 1767) известна с олигоцена (Петров, 1966, Зенкевич, 1977, Наумов, 2006 и др.), в Арктике - с миоцена. В морях арктического бассейна отмечен на глубинах 5–30 и 80–220 м (реже 0–5 м, 30–80 м и 220–1290 м) при температуре от - 2,0 до 13,7°С и солености 23,9 - 35,5‰, в основном на илистых (34% находок) и смешанных (40% находок) грунтах. На глубинах от 30 до 80 м – преобладает Hiatella pholadis (Linnaeus, 1771). Широко распространенный субтропическо-арктический циркумполярный вид отмечен во всех морях Северного Ледовитого океана. Массовые находки этого вида с толстыми раковинами почти прямоугольных очертаний в моновидовых и обедененных комплексах часто говорят о холодноводных условиях обитания, часто рядом с придонной областью ледников, спускающихся в море (в отложениях плейстоцена фиорда Уддевалла, Южная Швеция), п-ов Литке (Северный остров, арх. Новая Земля), в бореальных комплексах это вид также встречается, но обычно не доминирует и характеризует более тонкостенными раковинами округлых очертаний. Раковины хиателл являются одним из лучших раковин морских моллюсков для абсолютного датирования. Сохранность. Раковины и обломки раковин тонкостенные, иногда - темного цвета, неокатанные или слабоокатанные, плотные, средних и небольших размеров со следами первичной окраски. Возраст. Миоцен-голоцен. Из-за относительно слабой фоссилизации, неоплейстоцен. Наиболее близкий комплекс встречен в среднем неоплейстоцене (эпоха похолодания) (Крылов, Гусев, 2010). Условия обитания. Вероятно, песчаные и глинистые илы шельфа Арктического бассейна. Наличие раковин морских моллюсков с двумя сомкнутыми створками может свидетельствовать об их возможном захоронении на месте обитания и морском генезисе вмещающих их осадков, из которых была отобрана данная проба. Образец № 3298/15 Образец № 3298/17 Букцинумы – бентосные обитатели арктических морей. Обитают как на мягких глинистых, песчано-глинистых и песчанистых так и твердых каменистых грунтах на глубинах 5–300 м (редко более) при солености 25,0–35,5‰ и низких -2–12oС температурах (Петров, 1966, Наумов, 2006). Natica clausa (Broderip et Sowerby, 1829) - бореально-арктический вид обитает в Атлантическом океане на юг до мыса Гаттерас по Американскому и Португалии по Европейскому побережью, в западной части Средиземного моря, в Северном Ледовитом океане на западе до западной и восточной Гренландии, на северо-западе до Земли Франца-Иосифа и арх. Шпицберген в восточной части моря Лаптевых, в Тихом океане от Берингова пролива по азиатскому побережью на юг до северной части Японского моря, по американскому - на юг до о-ва Ванкувер от нижней литорали (5 м) до глубины 2660 м. преимущественно на илистых и илисто-песчаных грунтах, при температуре воды от -1,4 до 16oС и солености 25,0–35,5‰ (Петров, 1966, Наумов, 2006). Мии – обитатели мягких илистых или песчано – илистых или реже песчаных грунтов морей Северного Ледовитого океана и морей севера Атлантического и Тихого океанов в миоцене-голоцене (https://en.wikipedia.org/wiki/Mya truncata # /media/File:Mya-truncata_in-situ_hg.jpg). Сохранность. Раковина и обломки раковин тонкостенные, обелены, плотный, мелованные, неокатанные, легкие, небольших размеров, слабофоссилизированые иногда со следами первичной окраски. Возраст. Миоцен-голоцен. Из-за относительно слабой фоссилизации, вероятно, неоплейстоцен, наличие букцинума говорит о возможности принадлежности этого комплекса к эпохе потепления (Крылов, Пенделяк, Костин, 2019). Более точное определение возраста не представляется возможным. Условия обитания. Вероятно, песчаные и глинистые илы шельфа Арктического бассейна. Фотографии коллекции изученных морских моллюсков Северной Земли (о. Большевик), сборы 2020 г. Фото 1–2. Четыре раковины морских двустворчатых моллюсков Mya truncata Linnaeus, 1758 из пробы 3298/7 (с двумя сомкнутыми створками и конхиолинновым слоем), которые свидетельствуют о прижизненном захоронении (вид сверху и снизу). Длина нижней правой раковины 2,5 см Фото 3–4. Четыре раковины морских двустворчатых моллюсков Mya truncata Linnaeus, 1758 из пробы 3298/7 (с двумя сомкнутыми створками и конхиолинновым слоем), которые свидетельствуют о прижизненном захоронении (вид слева и справа). Ширина левой раковины 1,2 см Фото 5. Многочисленные поврежденные раковины морских двустворчатых моллюсков Mya truncata Linnaeus, 1758 и их обломки из пробы 3298/7 и конхиолинновым слоем (вид сверху). Длина поврежденной нижней левой раковины 1,5 см Фото 6. Многочисленные поврежденные раковины морских двустворчатых моллюсков Mya truncata Linnaeus, 1758 и их обломки из пробы 3298/7 (вид снизу). Длина поврежденной нижней левой раковины 1,5 см Фото 7–8. Раковины морских моллюсков Mya truncata Linnaeus, 1758 и Hiatella arctica (Linaneus, 1758) из пробы № 3298/15. Длина левой раковины 3 см Фото 9–10. Раковина морского моллюска Natica clausa (Broderip et Sowerby, 1824) со следами первичной окраски и обломки раковин морских моллюсков Natica clausa (Broderip et Sowerby, 1829), Mya sp., Buccinum sp. из пробы № 3298/17. Диаметр левой раковины 1 см Фото 11. Раковина и обломок раковины морского моллюска Natica clausa (Broderip et Sowerby, 1829) со следами первичной окраски Фото 12. Внешний вид обнажения 1, из которого происходят изученные раковины моллюсков. Остров Большевик, архипелаг Северная Земля. Фото Д. Ю. Большиянова, 2020 г Фото 13–14. Внешний вид обнажения 2 с захороненными раковинами мий с двумя сомкнутыми створками (в прижизненном положении) и сифонами вверх, что свидетельствует о морском происхождении вмещающих их пород. Остров Большевик, архипелаг Северная Земля. Фото Д. Ю. Большиянова, 2020 г













ФАУНА МОРСКИХ МЛЕКОПИТАЮЩИХ
Рис. 1 Сборы костей морских млекопитающих, вымытых из отложений обнажения 3298 
ДАТИРОВАНИЕ ОТЛОЖЕНИЙ
Радиоуглеродное датирование выполнено в Лаборатории геоморфологических и палеогеографических исследований полярных регионов и Мирового океана им. Кёппена СПбГУ. Датирован ствол древесины плавника из пачки песчано-гравийных морских осадков, перекрывающих глины. Радиоуглеродный возраст образца составил 45760±1160 лет (календарный возраст 48630±1640 лет).
КРАТКАЯ НАУЧНАЯ ИНТЕРПРЕТАЦИЯ
ЛИТЕРАТУРА