
п-ов Канин
 |
 |
 |
Широта: 67°12' с.ш. Площадь: 5,8 км2 Площадь водосбора: 38,6 км2 |
{spoiler title=ХАРАКТЕРИСТИКА РАЙОНА и ОБЪЕКТА ИССЛЕДОВАНИЯ opened=0}
Озеро Каменное расположено в 9,5 км к востоку от мыса Конушин западного побережья п-ова Канин. Оно является остаточным водоёмом некогда большего озера, развивавшегося к югу от Шемоховских сопок. На склонах котловины современного озера есть террасы высотой 1,5 и 4 м. Мелкосопочный рельеф и термокарстовые впадины озёр свидетельствуют о таянии мёртвых льдов как причине формирования местных озёр. Озеро находится на высоте 19 м над уровнем моря. Котловина озера состоит из двух основных впадин - 6,5 и 5,3 м, максимальная обнаруженная глубина 7,7 м. Сток из озера осуществляется на юг по р. Каменной. Озеро располагается в подзоне типичной кустарничковой тундры, чуть севернее границы с лесотундрой.
{/spoiler}
{spoiler title=ИЗУЧЕННОСТЬ и ФАКТИЧЕСКИЙ МАТЕРИАЛ opened=0}
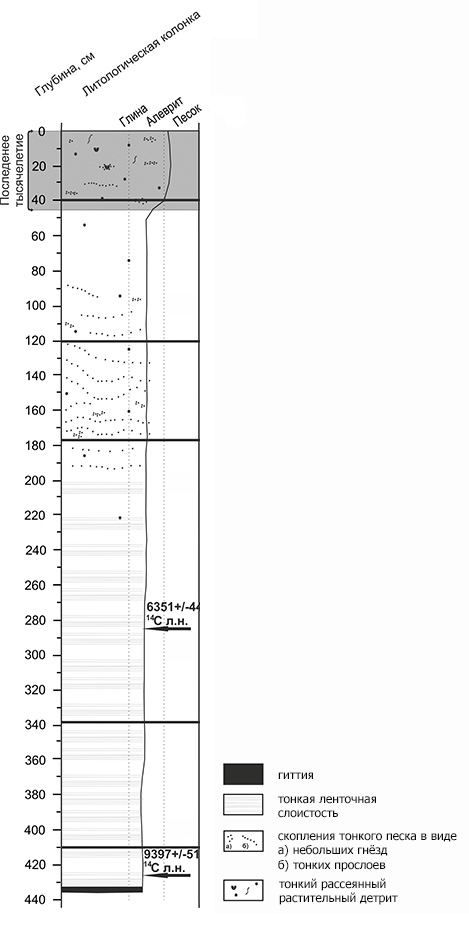
Керны длинной 4,3 м и 4 м были получены с глубины 4,3 и 2 м соответственно. Выше 2 метров осадок насыщен органическим детритом и отличается прекрасной горизонтальной слоистостью глин с тонкими черными прослоями органики.
 |
Фрагмент керна из оз. Каменное |
|
|
Слои гиттии в основании колонки, извлеченной из оз. Каменное |

В основании одного из кернов обнаружены слои гиттии (высокоорганогенного ила). Наблюдаемая мощность этого слоя 3 см. Накопление его происходило в очень благоприятных для развития растительности условиях, возможно, в климатический оптимум голоцена. В настоящее время этот регион расположен в пределах типичной тундры с бедной кустарничковой растительности.
| № керна | Координаты | Характеристика керна | глубина отбора, м | мощность, см | виды анализов |
| К-К-1 | 67°12,063'с.ш., 44°02,986'в.д. |
В основании керна обнаружены слои гиттии (высокоорганогенного ила). Наблюдаемая мощность этого слоя 3 см. |
4,3 | 436 |
|
{/spoiler}
{spoiler title=ЛИТОЛОГИЯ, ВАРВОХРОНОЛОГИЯ, СКОРОСТЬ ОСАДКОНАКОПЛЕНИЯ и ГРАНУЛОМЕТРИЯ opened=0}

{/spoiler}
{spoiler title=ГЕОХИМИЧЕСКИЙ АНАЛИЗ opened=0}
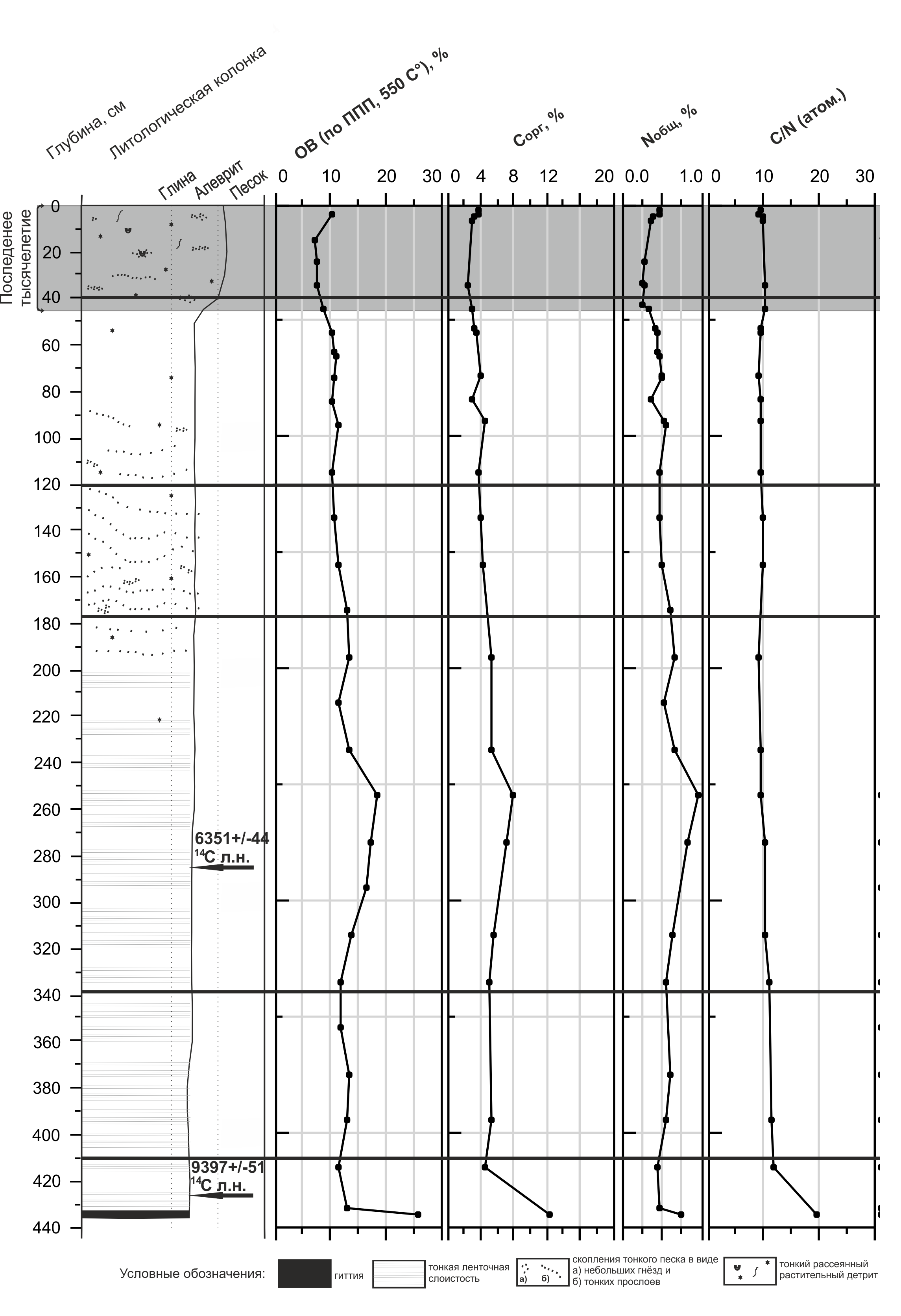
Геохимия донных отложений озера Каменного (Кольский п-ов)
{/spoiler}
{spoiler title=СПОРОВО-ПЫЛЬЦЕВОЙ АНАЛИЗ opened=0}
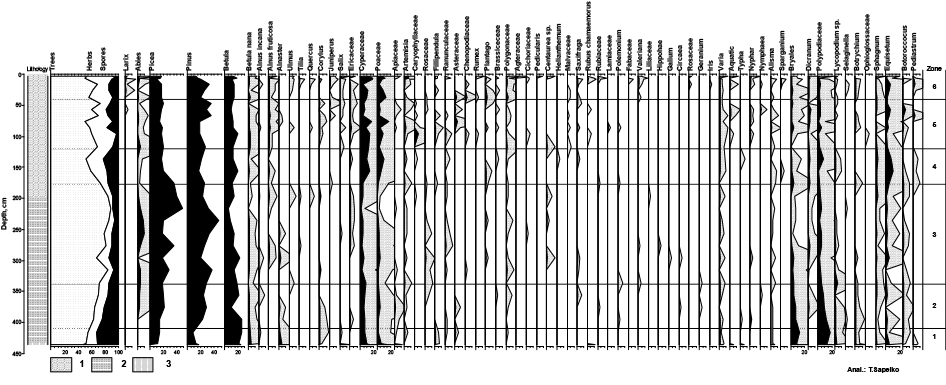
к.г.н. Сапелко Т.В.
Институт озероведения РАН
Предварительные результаты палинологического анализа донных отложений оз. Каменного
Из колонки донных отложений оз. Каменного мощностью 436 см просмотрено 30 образцов на спорово-пыльцевой анализ. Техническая обработка проводилась по стандартной методике (Гричук, 1940; Berglund, B. E. & Ralska-Jasiewiczowa, 1986) с применением тяжелой жидкости (CdI2 + KI). По результатам анализа построена спорово-пыльцевая диаграмма с помощью компьютерных программ TILIA2, TILIA GRAPH2 (Grimm, 1991) и TGView, на которой выделено 6 зон.
Палинозона 1 (436-410 см)
В основании разреза выделены спектры, хорошо насыщенные пыльцой. Среди пыльцы древесных пород (до 55%) преобладают береза и хвойные породы. Доминирует пыльца березы (до 25%). Споры также составляют значительный процент (21-32%). Преобладают Polypodiaceae и Bryales. Проценное содержание пыльцы трав относительно невелико (12-30%).
Площадь лесов значительна. В основном это березовые леса. Роль открытых травянистых сообществ менее значима. В основном преобладают осоковые и злаки. Разнообразие травянистого покрова выражено слабо. При этом в самом начале зоны распространяются водные и прибрежно-водные растения. Отмечен даже термофильный вид Typha latifolia. В это же время отмечены водоросли Pediastrum и Botryococcus.
Палинозона 2 (410-338 см)
Далее выделенная зона отличается увеличением пыльцы древесных (до 70%). Доминируют хвойные породы. Помимо увеличения количества пыльцы сосны и ели, появляется пыльца пихты. К концу зоны наряду с пихтой появляется ольха. Среди трав доминирует пыльца Cyperaceae. Значительно участие пыльцы злаков, полыни. Видовой состав трав становится более разнообразным. Среди спор доминанты в основном не меняются. Постоянно встречаются водоросли Botryococcus, а Pediastrum исчезают совсем.
Площадь лесов расширяется. Меняется доминанта. Распространяются хвойные леса при сокращении роли березы в лесных сообществах. Немного увеличивается разнообразие травянистого покрова.
Палинозона 3 (338-177 см)
Далее количество пыльцы древесных пород еще больше увеличивается и достигает своих максимальных значений (до 88%). Вновь меняется состав лесов. Среди пыльцы древесных появляются широколиственные породы (Ulmus, Quercus, Tilia). Доминирует пыльца сосны.
Среди пыльцы трав преобладает пыльца Cyperaceae. Постоянно встречается пыльца Rosaceae, Ranunculaceae, Polygonaceae. Ксерофиты практически исчезают. Распространяются луговые сообщества. Количество пыльцы водных растений невелико.
В это время отмечаются наиболее благоприятные условия для произрастания растительного покрова. Климат становится теплым и влажным. Возможно, в район озера продвинулась среднетаежная зона. Говорить о южнотаежной зоне, видимо нельзя, т.к. несмотря на обилие широколиственных пород, преобладают хвойные.
Палинозона 4 (177-120 см)
Уменьшается количество пыльцы древесных пород (50-73%). Процентный состав пыльцы трав достигает 25%. Среди древесных доминирует пыльца сосны и ели. Исчезает пыльца пихты, широколиственных пород. Среди трав доминирует пыльца злаков и осоковых. Видовое разнообразие несколько снижается. Увеличивается количество и разнообразие спор. Вновь появляются водоросли Pediastrum. Здесь они отмечены двух видов.
Полностью исчезают широколиственные породы из состава лесов.
Площади открытых пространств немного увеличиваются. Климат, видимо, становится более холодным и сухим.
Палинозона 5 (120-40 см)
Количество пыльцы древесных пород остается примерно без изменений. Процент пыльцы трав возрастает и достигает здесь своего максимума по разрезу (до 41%). Состав лесов вновь меняется. Вновь появляется и постоянно присутствует пыльца пихты. К концу зоны появляется пыльца лиственницы. В середине зоны единично встречается одновременно пыльца вяза и дуба. Доминирует при этом все также пыльца сосны и ели.
Состав трав в этот период достигает максимального разнообразия.
Преобладает пыльца Cyperaceae, Poaceae, Ericaceae, Polygonaceae. Определена пыльца жимолости (Lonicera xylosteum), характерной для хвойных и смешанных северных лесов. Резко возрастает количество и разнообразие пыльцы водных растений. Растет также разнообразие спор.
Климат, видимо, становится несколько суше и теплее. Разнообразие лесных ассоциаций говорит о некотором восстановлении благоприятных для растительности условий. Наряду с лесными и луговыми травами появляются открытые сообщества с ксерофитами. Очень высокое содержание водных и прибрежно-водных растений может свидетельствовать о некотором зарастании водоема. В целом можно говорить о большом разнообразии растительного покрова.
Первый выделенный период (зона 1) сформировался в достаточно прохладном и в меру влажном климате, способствующему развитию березовых и хвойных лесов. При этом наибольшее распространение в это время получили березовые леса. Выводы палинологического заключения о пребореальном возрасте этой зоны подтвердила полученная радиоуглеродная датировка 9397+51 (гл. 431-423 см). Далее (зона 2) накапливались бореальные отложения. Широко распространяются хвойные леса, вытесняя березовые. Дальше можно говорить о наступлении наиболее благоприятных климатических условиях (зона 3) и отнесению этого периода к Атлантике, что подтвердила радиоуглеродная датировка 6351+44 (гл. 287-283 см). Доминируют хвойные леса с примесью широколиственных пород. Следующий выделенный период (зона 4) отличался темнохвойными сомкнутыми лесами. Датировать его можно суббореальным периодом. Небольшое похолодание, возможно, соответствует началу суббореального периода (SB-1). Дальнейшее потепление (зона 5) можно соотнести с серединой суббореала (SB-2) и суббореальным климатическим оптимумом, выделенном Н.А. Хотинским (1977) и отмечаемом во многих разрезах северных территорий.
Последний описанный период (зона 6), возможно, отнести к завершающему этапу суббореального периода (SB-3). Ранее мы предполагали субатлантический возраст верхней зоны. Однако с большой натяжкой ее можно отнести к современному периоду. Несколько смущает активное развитие водных и прибрежно-водных растений, наличие термофильных видов. При этом общая концентрация пыльцы довольно низкая относительно предыдущих этапов.
{/spoiler}
{spoiler title=ДИАТОМОВЫЙ АНАЛИЗ opened=0}
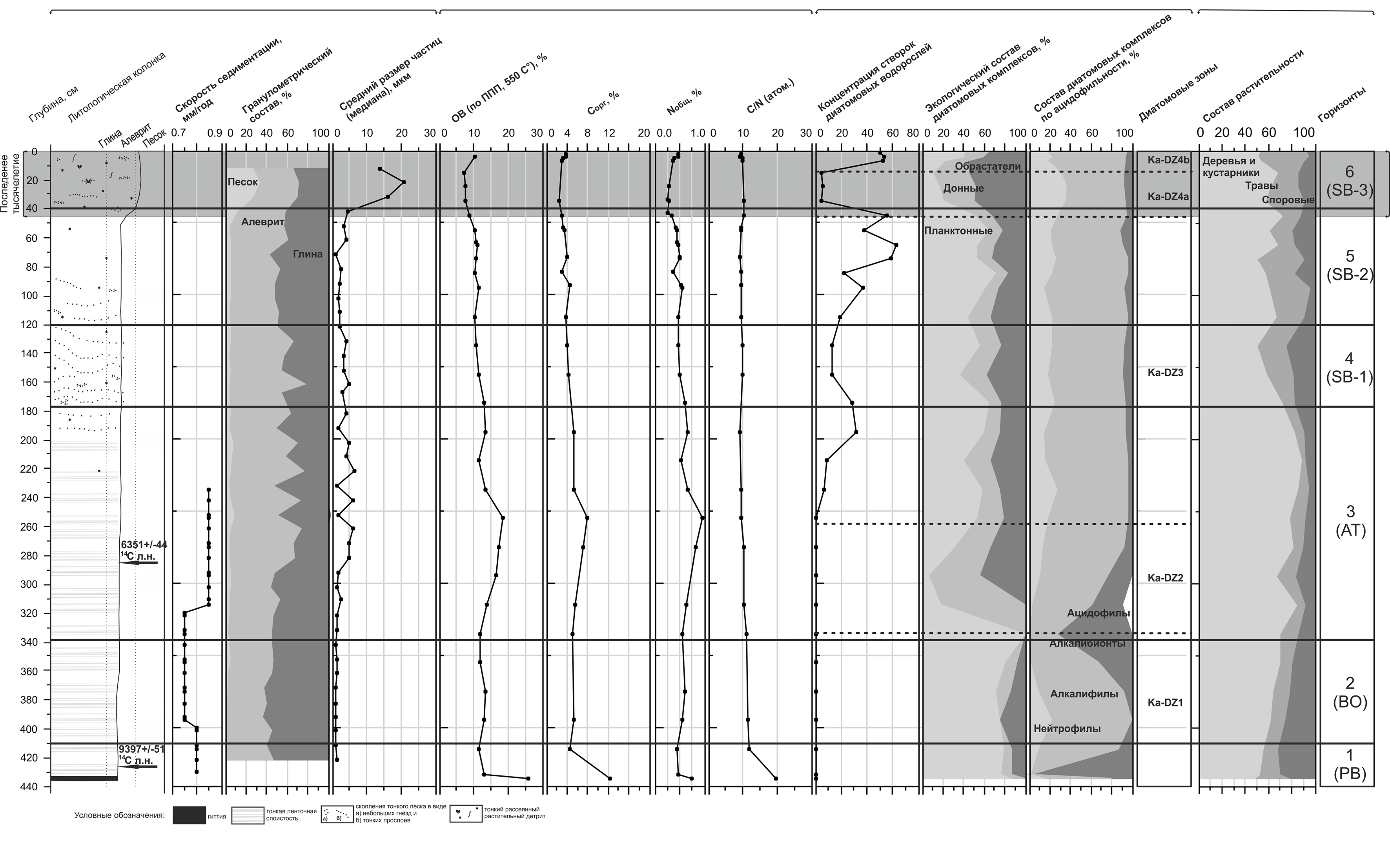
Диатомовая диаграмма озера Каменного: относительная численность основных видов диатомей, концентрация створок (млн) в 1 г сухого осадка, соотношение основных экологических групп и диатомовые зоны (Палеолимнология…, 2008).

Лудикова А.
Институт озероведения РАН, г. Санкт-Петербург
Мощность исследованной колонки донных осадков составила 436 см. Донные отложения имеют следующий состав: 0-200 см – алеврит, насыщенный растительным детритом, 200-433 см – глинистый алеврит с прослоями органики, 433-436 см – торф с древесными остатками.
Диатомовый анализ донных отложений оз. Каменное позволил выделить 4 диатомовые зоны.
Диатомовая зона Ka-DZ1 (435-333 см).
Данная зона характеризуется, в первую очередь, крайне низкими значениями концентрации створок диатомовых водорослей. Отмечены единичные панцири планктонных Aulacoseira ambigua (Grun.) Sim., A. islandica (O. Müll.) Sim., A. subarctica (O. Müll.) Haworth, Cyclostephanos dubius (Fricke) Round, обрастателей Achnanthes lanceolata (Bréb.) Grun., A. oestrupii (A. Cl.) Hust., Cymbella sinuata Greg., Fragilaria construens (Ehr.) Grun. et var. и Opephora martyi Hérib., а также донных диатомей рода Navicula (N. jentzschii Grun., N. scutelloides W. Sm.). По отношению к рН большинство встреченных здесь видов является алкалифилами, т.е. предпочитает щелочную среду обитания, менее многочисленны алкалибионты и нейтрофилы. Aulacoseira islandica и A. subarctica, створки которых преобладают в осадках данной зоны, являются типичными представителями диатомового планктона холодноводных озер (Прошкина-Лавренко, 1974).
Помимо пресноводных видов, в составе диатомовых комплексов зоны Ka-DZ1 отмечены также фрагменты створок морских диатомей Paralia sulcata (Ehr.) Sim., Stephanоpyxis spp. и др.
Диатомовая зона Ka-DZ2 (333-254 см).
Концентрация створок диатомовых водорослей немного увеличивается, при этом отмечается резкое уменьшение содержания планктонных видов и рост численности донных диатомей и обрастателейю В направлении верхней границы зоны численность донных и эпифитных таксонов вновь снижается, в то время как содержание створок планктонных диатомей значительно возрастает, в первую очередь за счет Aulacoseira islandica, достигающей здесь 30%.
Наиболее типичными среди обрастателей являются северо-альпийский Achnanthes oestrupii, бореальные Cocconeis disculus (Schum.) Cl. и C. neodiminuta Krammer, а также имеющие широкое географическое распространение виды-космополиты F. brevistriata Grun., F. construens et var. и Opephora martyi. Донные диатомеи представлены видами Gyrosigma attenuatum (Kütz.) Rabenh., Navicula aboensis (Cl.) Hust., N. jentzschii и N. scutelloides – характерными обитателями литоральной зоны холодноводных олиготрофных бассейнов (Давыдова, 1985; Hedenström & Risberg, 1999).
Отмечается наибольшая за всю исследованную историю озера численность алкалибионтов – организмов, развивающихся исключительно в щелочных водах, представленных, главным образом, видами G. attenuatum и N. scutelloides.
Диатомовая зона Ka-DZ3 (254-45 cм).
В интервале 254-195 см отмечается существенное возрастание концентрации створок диатомовых водорослей. Вверх по разрезу ее значения несколько снижаются, вновь возрастая в интервале 85-45 см, где они достигают максимальных значений за всю историю озера.
Доминируют планктонные диатомеи, численность которых, как правило, превышает 50%. Как и в предыдущих зонах, здесь доминируют Aulacoseira ambigua, A. islandica и A. subarctica, а Cyclostephanos dubius приобретает значение субдоминанта.
Среди эпифитов преобладают представители рода Fragilaria (F. construens et var., F. exigua Grun., F. pinnata Ehr.). Происходит небольшое увеличение относительного содержания Achnanthes lanceolata et f. frequentissima Lange-B., A. oestrupii и Cymbella sinuata, в составе диатомовых комплексов появляется также северо-альпийский A. calcar Cl.. Донные диатомеи представлены, главным образом, видами рода Navicula, среди которых наиболее многочисленны бореальные N. jaernefeltii Hust. и N. jentzschii. Необходимо отметить значительное сокращение числа видов-алкалибионтов по сравнению с предыдущей зоной.
Зона Ka-DZ4 (45-0 см), подзона Ka-DZ4а (45-15 см).
Подзона характеризуется резким снижением концентрации створок диатомовых в донных отложениях. Сокращается процент планктонных диатомей, возрастает численность эпифитов и обитателей донного субстрата, суммарное содержание которых составляет более 80%.
В число обрастателей, помимо Fragilaria pinnata, F. construens et var. и диатомей рода Achnanthes, в рассматриваемом интервале входит также вид Opephora martyi, до сих пор крайне редко встречавшийся в составе диатомовых комплексов.
По сравнению с предыдущей зоной, отмечается рост относительного содержания донных Navicula jentzschii и N. aboensis.
Подзона Ka-DZ4b (15-0 см).
Концентрация створок диатомей скачкообразно возрастает.
Отмечается рост относительного содержания планктонных диатомей до 40%, представленных, в основном, видами Aulacoseira ambigua, A islandica, A subarctica и Cyclostephanos dubius. Численность обрастателей и донных диатомей снижается, однако, видовой состав доминантов в этих экологических группах остается тем же, что и в подзоне Ka-DZ4а.
Выделяемые по отношению к рН среды виды-алкалифилы по-прежнему численно преобладают. Процент диатомей-нейтрофилов сокращается почти вдвое по сравнению с предыдущей подзоной.
По результатам диатомового анализа донных отложений озера Каменного можно сделать вывод о том, что на протяжении всей истории в его котловине сохранялись пресноводные, преимущественно слабощелочные условия. В составе диатомовых комплексов преобладают северо-альпийские и бореальные диатомеи, характерные для водоемов арктических и умеренных широт.
Тот факт, что в прослое торфа, обнаруженном в подошве разреза донных отложений, отсутствуют диатомеи, как правило, в массе развивающихся в ходе заболачивания и заторфовывания водоемов, свидетельствует о том, что его образование происходило в субаэральных условиях при дефиците влаги. Планктонные виды, обнаруженные в этом горизонте, типичны для диатомовых комплексов последующих этапов развития озера и, по всей видимости, попали в торф из вышележащей толщи. Таким образом, начало аккумуляции перекрывающих торфянистый горизонт озерных осадков в котловине озера Каменного, по всей вероятности, можно считать временем образования данного водоема.
Находки морских диатомей не дают основания для предположения о проникновении морских вод в котловину озера даже на самых ранних этапах развития. Их плохая сохранность, а главное – отсутствие в соответствующих горизонтах солоноватоводных видов, массово развивающихся при переходе от морских условий к пресноводным (напр., Corner et al., 2001), указывают на несостоятельность подобного предположения. Кроме того, оно явно противоречит имеющимся палеогеографическим данным о позднечетвертичной истории региона, согласно которым основная часть территории п-ова Канин, включая район озера Каменного, последний раз подвергалась оледенению 55-45 тыс. лет назад (Larsen et al., 2006).
Во время максимума последнего, Валдайского оледенения край ледника располагался на расстоянии 15-25 км западнее п-ова Канин (Demidov et al., 2006). Следовательно, район исследования не испытывал ледниковой нагрузки и не мог быть затоплен морскими водами в ходе эвстатического подъема уровня Мирового океана. Таким образом, единственным объяснением присутствия в составе диатомовых комплексов озера Каменного экологически несовместимых пресноводных и морских видов является переотложение последних из более древних, микулинских морских осадков, отмеченных в многочисленных разрезах п-ова Канин (Kjær et al., 2006).
Поступление в озеро створок морских диатомей на начальном этапе развития (диатомовая зона Ka-DZ1) могло происходить в ходе вымывания их из указанных морских осадочных толщ, связанного с улучшением климатических условий и, возможно, таянием многолетней мерзлоты. Доминирование в составе диатомовых комплексов планктонных видов свидетельствует об обстановках, благоприятствующих развитию этой экологической группы: уровень озера на рассматриваемой стадии, по всей видимости, повышался. С другой стороны, активизация эрозионных процессов, по-видимому, имела своим результатом увеличение роли терригенного разбавления и рост темпов седиментации, о чем свидетельствуют крайне низкие значения концентрации створок диатомей. Увеличение объемов поступления аллохтонного вещества в котловину озера должно было привести к снижению прозрачности водной массы, что затрудняло проникновение солнечной радиации сквозь толщу воды и тем самым препятствовало развитию диатомей бентоса. В такой обстановке фитопланктон, осуществляющий фотосинтез в верхней части водной толщи, оказывался более «конкурентоспособным». С другой стороны, незначительное содержание панцирей диатомовых водорослей в донных отложениях может также объясняться низкой продуктивностью водоема на рассматриваемой стадии.
Последовавшее затем увеличение численности донных диатомей (интервал 334-294 см, диатомовая зона Ka-DZ2) могло стать следствием понижения уровня водоема, возможно, в связи с образованием стока, либо некоторого похолодания климата и уменьшения количества атмосферных осадков. В результате возросла площадь мелководий, пригодных для колонизации бентосными диатомовыми водорослями. О широком распространении крайне мелководных условий свидетельствует, в частности, присутствие аэрофильной диатомеи Ellerbeckia arenaria f. teres (Brun) Crawford, достигающей в рассматриваемой зоне наиболее высокой численности за всю историю водоема. Кроме того, похолодание могло привести к увлечению продолжительности периода ледостава и сокращению сроков вегетационного периода, что, в свою очередь, также не способствовало развитию планктонной диатомовой флоры (Smol, Cumming, 2000).
Тот факт, что в составе диатомовых комплексов на данном этапе продолжают встречаться переотложенные створки морских диатомей, по-видимому, также свидетельствует об уменьшении глубины озера на данном этапе: понижение уровня водоема, вероятно, привело к абразии морских отложений Микулинского межледниковья.
Значительное увеличение доли диатомей-алкалибионтов на рассматриваемом этапе представляется трудно объяснимым исходя лишь из результатов диатомового анализа, поскольку в периоды похолоданий рН водной среды, как правило, уменьшается (напр., Smol, Cumming 2000).
В целом продуктивность водоема в указанный период оставалась достаточно низкой, на что, очевидно, указывают низкие значения концентрации створок диатомей в донных осадках.
Рост содержания планктонных диатомей, отмечаемый на следующем этапе развития (диатомовая зона Ka-DZ3), скорее всего, явился результатом очередного увеличения глубины водоема, что, в свою очередь, могло быть связано с потеплением и увлажнением климата.
Доминирующие здесь Aulacoseira islandica и A. subarctica являются типичными представителями диатомового планктона северных озер, в частности, озер Большеземельской тундры, Кольского полуострова и др. (Большие озера…, 1975, Особенности структуры…, 1994). Однако преобладание в диатомовых комплексах зоны Ka-DZ3 этих видов, обычно относимых к так называемым северо-альпийским, в данном случае указывает не на ухудшение климатических условий, а, напротив, на потепление и/или увлажнение климата и повышение содержания биогенов, в первую очередь, растворенного фосфора и кремнезема.
Кроме того, высокая численность представителей рода Aulacoseira может говорить также об увеличении глубины, так как вследствие массивности кремневых панцирей большинство из них, как правило, нуждается в определенной мощности водной толщи.
О некотором росте обеспеченности биогенными элементами в рассматриваемый период свидетельствует присутствие в составе диатомовых комплексов вида Cyclostephanos dubius, как правило, обитающего в высокотрофных условиях. По-видимому, поступление минеральных веществ в этот период возросло в связи с увеличением поверхностного стока. В то же время в литоральной зоне обитают такие типичные виды олиготрофных северных водоемов, как Achnanthes oestrupii, Navicula jaernefeltii и N. jentzschii, что вероятно, указывает на в целом невысокий уровень трофности.
Улучшение климатической обстановки сказалось на повышении продуктивности экосистемы водоема в целом, что нашло отражение в увеличении концентрации створок диатомей.
Новый этап развития озера Каменного, характеризующийся резким изменением соотношения основных экологических групп диатомей и общим снижением продуктивности водоема, по-видимому, был обусловлен очередной сменой природно-климатических обстановок. Снижение численности планктонных видов, как и в интервале, соответствующем диатомовой зоне Ka-DZ2, вероятно, явилось результатом некоторого обмеления озера в связи с уменьшением количества атмосферных осадков и/или общим похолоданием. Сокращение поступления биогенных элементов с поверхностным стоком, очевидно, стало причиной заметного снижения содержания видов Aulacoseira ambigua и Cyclostephanos dubius, предпочитающих условия более высокой трофности. В свою очередь, развитию обрастателей, в частности, Fragilaria pinnata могли способствовать обстановки более короткого безледного периода, продолжительность которого на озерах Кольского полуострова и Большеземельской тундры в настоящее время составляет 2-3(4) месяца. Поскольку F. pinnata хорошо приспособлена к условиям недостаточной освещенности, связанным в арктических озерах с более длительным существованием ледового покрова, ее численность может возрастать в периоды похолоданий (Westover et al., 2006).
В целом же скачкообразный характер изменений, зафиксированных по результатам диатомового анализа, делает их интерпретацию и реконструкцию палеогеографической обстановки чрезвычайно затруднительными без привлечения результатов других исследований.
Для позднейшей историй озера Каменного вновь характерен рост продуктивности и установление условий, более благоприятных для развития диатомей планктона, что соответствует современному состоянию этого водоема.
В заключении необходимо подчеркнуть, что интерпретация полученных данных не претендует на полноту и точность, и настоятельно требует подтверждения датировками и результатами других анализов, в первую очередь, спорово-пыльцевого и геохимического (Палеолимнология…, 2008).
{/spoiler}
{spoiler title=РАДИОУГЛЕРОДНОЕ ДАТИРОВАНИЕ opened=0}
 |
Радиоуглеродное датирование проведено в Christian-Albrechts-University, Kiel, Germany.
|
{/spoiler}
{spoiler title=КРАТКАЯ НАУЧНАЯ ИНТЕРПРЕТАЦИЯ opened=0}
Результаты анализов колонки донных отложений К-К-1, оз. Каменное, п-в Канин (Большиянов и др., 2011)
Палинозона 1 (436-410 см). Доминируют древесные породы (до 55%), среди которых преобладает береза (до 25%) и хвойные. Споры составляют 21-32%, пыльца трав – 12-30% с преобладанием осок и злаков. Вероятно, берёзовые леса занимали значительные площади с небольшой ролью открытых травяных сообществ.
Климат прохладный и в меру влажный. Зона отнесена к пребореальному периоду, что подтверждено радиоуглеродной датировкой 9395+50 (KIA38586) мелкой органики с глубины 431-423см. Отложения представлены алевритово-глиняными осадками, местами с тонкой неясной горизонтальной слойчатостью (вероятно годовых циклов ленточных глин). В подошве керна слой гиттии (436-433 см). Содержание органического вещества (ОВ) в нём составляет около 25% (ППП≈25%, Сорг ≈12%), выше резко падает – ППП≈11-13%, Сорг ≈5%. Отношение C/N соответственно уменьшается с 20 до 11, что указывает на изменение источника ОВ. Для гиттии им являлась водная и прибрежно-водная растительность, в вышележащих осадках органика имеет преимущественно аутигенную природу. Пыльца прибрежно-водной растительности массово распространяется в начале зоны. Среди диатомовых водорослей преобладают планктонные виды, но концентрация створок в осадке низкая, что можно объяснить значительным терригенным разбавлением. Большинство видов диатомовых является алкалифилами, менее многочисленны алкалибионты и нейтрофилы, что указывает на слабощелочную реакцию среды. В основном это представители холодноводных олиготрофных озёр. Отмечены также фрагменты створок морских диатомей – Paralia sulcata (Ehr.) Sim., Stephanоpyxis spp. и др., плохой сохранности.
Палинозона 2 (410-338 см) отличается увеличением пыльцы древесных пород (до 70%) с преобладанием хвойных: сосны и ели. Отмечается пыльца пихты, к концу зоны – ольхи. Доминанты среди трав и споровых растений в основном не меняются, но состав травянистого покрова становится богаче. Площадь лесов расширяется, при этом начинают доминировать хвойные породы за счёт сокращения роли берёзы. Выделенная зона соотнесена с бореальным периодом голоцена. Другие показатели отложений существенно не меняются.
Палинозона 3 (338-177 см) охватывает интервал, в котором количество пыльцы древесных пород достигает максимальных значений (до 88%). В составе спектров доминирует пыльца сосны, появляются широколиственные породы: вяз, дуб, липа. Видовой состав трав обогащается, ксерофиты почти исчезают.
Климат интерпретируется как тёплый и влажный. Возможно, в район озера продвинулась среднетаёжная подзона с преобладанием хвойных пород деревьев. Выделенный интервал сопоставлен с атлантическим периодом, что согласуется с радиоуглеродной датировкой – 6350+45 (KIA 38585) c глубины 287-283 см. В Впервую половину этого периода (333-254 см) преобладают донные и эпифитные, северо-альпийские и бореальные виды диатомей, но общая концентрация створок продолжает оставаться низкой. Литологически отложения почти не изменились. Во второй половине зоны (254-195 см) возросла концентрация створок диатомей, что является отражением повышения продуктивности экосистемы водоёма. Преобладание планктонных видов, вероятно, свидетельствует о постепенном увеличении глубины водоёма, что подтверждается и высокой численностью представителей рода Aulacoseira. Трофность озера также повышается, поскольку растёт значение видов, требовательных к содержанию биогенных элементов (Aulacoseira ambigua, A. islandica и A. Subarctica, Cyclostephanos dubius). Изменения, связанные с глубиной и трофностью, обусловлены потеплением, увлажнением и увеличением стока в озеро, о чём говорят и литологические данные – в гранулометрическом составе отмечается небольшое увеличение доли песка (до 6-8%). Повышенные значения ППП≈16-18%, Сорг ≈6-8% в первой половине выделенной зоны достигают пика в момент скачка в концентрации створок диатомей. Затем эти показатели возвращаются на прежний уровень (ППП≈11-13%, Сорг ≈5%), хотя концентрация створок не падает. При этом значение C/N порядка 9-10 подчёркивает аутигенное происхождение органики. Ясной связи между содержанием в осадках створок диатомей, отражающей в некоторой степени продуктивность водоёма, и содержанием в них ОВ не установлено. В данном случае это не может быть объяснено поступлением органики извне. На аккумуляцию ОВ в осадках помимо первичной продукции планктона влияет множество других факторов и, поэтому, не всегда между ними есть линейная зависимость.
Палинозона 4 (177-120 см) фиксируется уменьшением значимости древесных пород (50-73%). Доминируют сосна и ель, а пыльца пихты и широколиственных пород почти исчезает; растёт содержание спор и пыльцы трав. Развиты темнохвойные сомкнутые леса, но площадь открытых пространств немного увеличивается из-за изменения климата в сторону более холодного и сухого. Зону можно отнести к началу суббореального периода (SB-1). Состав диатомовой флоры не изменяется, но концентрация створок водорослей заметно понижается. Постепенно меняется характер отложений – за счёт появления примеси рассеянного тонкого растительного детрита и волнистых текстур, слойчатость приобретает неясный характер и постепенно пропадает, т.е. гидродинамический режим становился более активным. Доля ОВ в осадках уменьшается (ППП≈11%, Сорг ≈4%), отношение C/N по-прежнему равно 9-10, так что поступавший терригенный материал, должно быть, в большей степени играет роль «разбавителя», нежели источника органического вещества.
Палинозона 5 (120-40 см). Количество пыльцы древесных пород остаётся на прежнем уровне, но меняется её состав за счёт присутствия пыльцы пихты, лиственницы, вяза, дуба. Содержание и разнообразие пыльцы трав возрастает и достигает максимума по разрезу (до 41%). Климат становился суше и теплее. Наряду с лесами и лугами существовали открытые сообщества с ксерофитами. Данное потепление можно соотнести с климатическим оптимумом середины суббореального периода (SB-2), выделенным Н.А. Хотинским и отмечаемым во многих разрезах северных территорий. В составе диатомей серьёзных изменений не происходит, но численность створок в осадках максимальна. Литологический тип отложений сохраняется, а содержание ОВ продолжает снижаться (ППП≈10-11%, Сорг ≈3-4%) на фоне прежнего отношения C/N.
Палинозона 6 (40-0 см). Среди древесных пород продолжают доминировать хвойные и берёза. Единична пыльца лиственницы и широколиственных пород. Среди кустарников есть пыльца можжевельника, ольхи, ивы. Это были леса северотаежного типа во влажных и прохладных условиях. С глубины 45 см резко увеличивается доля песчаной фракции в осадках. Одновременно с песком возрастает примесь растительного детрита. Усилившимся привносом грубого терригенного материала можно объяснить резкое снижения концентраций створок диатомей и пыльцы, а также величин ППП и Сорг (до 7-8 и 2-3%). Обилие пыльцы водной и прибрежно-водной растительности, изменения гранулометрического состава свидетельствует об обмелении и зарастании водоёма. Данный интервал, по спорово-пыльцевому составу, возможно, относится к завершающему этапу суббореального периода (SB-3), но т.к. верх колонки сохранился хорошо, он накапливался всё же в субатлантическое и современное время.
Расчёты, произведённые по данным радиоуглеродного датирования, дали величину скорости осадконакопления 0,45 мм/год.
Варвометрический анализ по отдельным отрезкам керна дал другие результаты. В целом среднее значение скорости осадконакопления составило 0,8 мм/год. (Большиянов и др., 2011)
{/spoiler}
{spoiler title=ЛИТЕРАТУРА opened=0}
Опубликованная литература
Berglund, B. E. & Ralska-Jasiewiczowa, M. Pollen analysis and pollen diagrams // Handbook of Holocene Palaeoecology and Palaeohydrology. 1986. P. 455-484.
Corner G.D., Kolka V.V., Yevzerov V.Y., Møller J. J. Postglacial relative sea-level change and stratigraphy of raised coastal basins on Kola Peninsula, northwest Russia // Global and Planetary Change. 2001, Vol. 31. p.155–177.
Demidov I.N., Houmark-Nielsen M., Kjær K.H., Larsen E. The last Scandinavian Ice Sheet in northwestern Russia: ice flow patterns and decay dynamics // Boreas. 2006. Vol. 35. p. 425-443.
Grimm E. Tilia 1.12, Tilia Graph 1.18. Illinois State Museum, Springfield, 1991.
Hedenström A., Risberg J. Early Holocene shore-displacement in southern central Sweden as recorded in elevated isolated basins // Boreas. 1999. Vol. 28. p. 490–504.
Kjær K. H., Larsen E., Funder S., Demidov I. N., Jensen M., Håkansson L., Murray A. Eurasian ice-sheet interaction in northwestern Russia throughout the late Quaternary // Boreas. 2006, Vol. 35. p. 444-475.
Larsen E., Kjær K.H., Demidov I.N., Funder S., Grøsfjeld K., Houmark-Nielsen M., Jensen M., Linge H., Lyså A. Late Pleistocene glacial and lake history of northwestern Russia // Boreas. 2006. Vol. 35, № 3. p. 394-424.
Smol J.P., Cumming B.F. Tracking long-term changes in climate using algal indicators in lake sediments // Journal of Phycology. 2000, Vol. 36. p. 986-1011.
Westover K., Fritz Sh., Blyakharchuk T., Wright H. Diatom paleolimnological record of Holocene climatic and environmental change in the Altai Mountains, Siberia // Journal of Paleolimnology. 2006, Vol. 35. p. 519-541.
Большие озера Кольского полуострова. Л.: Наука, 1976. 349 с.
Гричук В.П. Методика обработки осадочных пород, бедных органическими остатками, для целей пыльцевого анализа // Проблемы физической географии. Вып. 8,1940, с. 53-58.
Давыдова Н.Н. Диатомовые водоросли – индикаторы природных условий водоемов в голоцене. Л.: Наука, 1985. 244 с.
Даувальтер В.А. Закономерности распределения концентраций тяжелых металлов в донных отложениях в условиях загрязнения и закисления озер (на примере Кольского Севера) // Автореф. дисс. на соиск. учен. степ. канд. геогр. наук. – СПб, 1994.
Критерии оценки экологической обстановки территорий для выявления зон чрезвычайной экологической ситуации и зон экологического бедствия. (Утв. Минприроды РФ 30 ноября 1992 г.).
Лукин А.А., Даувальтер В.А., Новоселов А.П. Экосистема Печоры в современных условиях. – Апатиты: изд. КНЦ РАН, 2000.
Моисеенко Т.И. Теоретические основы нормирования антропогенных нагрузок на водоемы субарктики. – Апатиты: изд. КНЦ РАН, 1997.
Моисеенко Т.И., Даувальтер В.А., Родюшкин И.В. Геохимическая миграция элементов в субарктическом водоеме. – Апатиты: изд. КНЦ РАН, 1997.
Особенности структуры экосистем озер Крайнего Севера (на примере озер Большеземельской тундры). СПб.: Наука, 1994. 260 с.
Палеолимнология озера Каменного (п-ов Канин) по данным диатомового анализа донных отложений // Сборник научных трудов XII Всероссийской Палинологической конференции «Палинология: стратиграфия и геоэкология», Санкт-Петербург, 29 сентября – 4 октября, 2008 г., Т. I. С. 204-209.
Прошкина-Лавренко А.И. Диатомовые водоросли современных морей и озер // Диатомовые водоросли СССР. Л.: Наука, 1974. Т. 1. с. 274-351.
Хотинский Н. А. Голоцен современной Евразии, М.: Наука, 1977, 200 с.
{/spoiler}