
–Э–Њ–≤—Л–є, —А–∞–љ–µ–µ –љ–µ–Є–Ј–≤–µ—Б—В–љ—Л–є —А–∞–Ј—А–µ–Ј –Љ–Њ—А—Б–Ї–Є—Е —З–µ—В–≤–µ—А—В–Є—З–љ—Л—Е –Њ—В–ї–Њ–ґ–µ–љ–Є–є –Њ–±–љ–∞—А—Г–ґ–µ–љ –ї–µ—В–Њ–Љ 2020 –≥. –≤—Б–ї–µ–і—Б—В–≤–Є–µ —В–Њ–≥–Њ, —З—В–Њ –∞–љ–Њ–Љ–∞–ї—М–љ–Њ —В—С–њ–ї—Л–µ —Г—Б–ї–Њ–≤–Є—П –њ—А–Є–≤–µ–ї–Є –Ї –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ–Љ—Г —В–∞—П–љ–Є—О –ї–µ–і–љ–Є–Ї–Њ–≤, –Є —Б–љ–µ–ґ–љ–Є–Ї–Њ–≤, –≤ —З–∞—Б—В–љ–Њ—Б—В–Є –љ–∞ –Љ–∞—Б—Б–Є–≤–µ –≥–Њ—А—Л –Ч–≤—С–Ј–і–љ–Њ–є (416 –Љ –љ–∞–і —Г—А–Њ–≤–љ–µ–Љ –Љ–Њ—А—П), —А–∞—Б–њ–Њ–ї–Њ–ґ–µ–љ–љ–Њ–є –љ–∞–і –Ј–∞–њ–∞–і–љ—Л–Љ –њ–Њ–±–µ—А–µ–ґ—М–µ–Љ –Ј–∞–ї–Є–≤–∞ –Р—Е–Љ–∞—В–Њ–≤–∞ (–Њ. –С–Њ–ї—М—И–µ–≤–Є–Ї –∞—А—Е–Є–њ–µ–ї–∞–≥–∞ –°–µ–≤–µ—А–љ–∞—П –Ч–µ–Љ–ї—П). –Т –і–Њ–ї–Є–љ–µ –±–µ–Ј—Л–Љ—П–љ–љ–Њ–є —А–µ–Ї–Є, —Б—В–µ–Ї–∞—О—Й–µ–є —Б —Б–µ–≤–µ—А–љ—Л—Е —Б–Ї–ї–Њ–љ–Њ–≤ –≥–Њ—А—Л, –Є –≤–њ–∞–і–∞—О—Й–µ–є –≤ –Ј–∞–ї–Є–≤ –Р—Е–Љ–∞—В–Њ–≤–∞, –њ—А–Њ–Є—Б—Е–Њ–і–Є–ї–∞ –Є–љ—В–µ–љ—Б–Є–≤–љ–∞—П –±–Њ–Ї–Њ–≤–∞—П —Н—А–Њ–Ј–Є—П, –Ї–Њ—В–Њ—А–∞—П –Є –≤—Б–Ї—А—Л–ї–∞ —А–∞–љ–µ–µ –љ–∞—Е–Њ–і–Є–≤—И–Є–µ—Б—П –њ–Њ–і —И–ї–µ–є—Д–∞–Љ–Є –Њ—Б—Л–њ–µ–є –Љ–Њ—А—Б–Ї–Є–µ –Њ—В–ї–Њ–ґ–µ–љ–Є—П, –Ї–Њ—В–Њ—А—Л–µ –Њ–±–љ–∞–ґ–Є–ї–Є—Б—М –њ–Њ—З—В–Є –≤–µ—А—В–Є–Ї–∞–ї—М–љ—Л–Љ–Є —Б—В–µ–љ–Ї–∞–Љ–Є. –Ъ–∞–Ї –Є –±–Њ–ї—М—И–Є–љ—Б—В–≤–Њ –≤–Њ–і–Њ—В–Њ–Ї–Њ–≤ –њ–Њ –±–µ—А–µ–≥–∞–Љ –Ј–∞–ї–Є–≤–∞ –Р—Е–Љ–∞—В–Њ–≤–∞, –Ј–∞–Ї–∞–љ—З–Є–≤–∞—О—Й–Є–µ—Б—П –≤ –љ—С–Љ –ї–Њ–њ–∞—Б—В–љ—Л–Љ–Є –і–µ–ї—М—В–∞–Љ–Є, –Ї–Њ–љ—Г—Б –≤—Л–љ–Њ—Б–∞ —А–µ–Ї–Є, –Њ–±—А–∞–Ј–Њ–≤–∞–љ –Ї—А—Г–њ–љ–Њ–Њ–±–ї–Њ–Љ–Њ—З–љ—Л–Љ –Љ–∞—В–µ—А–Є–∞–ї–Њ–Љ, –∞–ї–µ–≤—А–Є—В–Њ–Љ –Є –≥—А–∞–≤–Є–µ–Љ, –≤—Л—Б—В—Г–њ–∞–µ—В –Ј–∞ –ї–Є–љ–Є—О –±–µ—А–µ–≥–∞ –љ–∞ 400 –Љ. –Ч–љ–∞—З–Є—В–µ–ї—М–љ–∞—П –Љ—Г—В–љ–Њ—Б—В—М –≤–Њ–і—Л –≤ —А—Г–Ї–∞–≤–∞—Е –і–µ–ї—М—В—Л –≤–Њ –≤—А–µ–Љ—П –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–є —Б–≤–Є–і–µ—В–µ–ї—М—Б—В–≤–Њ–≤–∞–ї–∞ –Њ–± –Є–љ—В–µ–љ—Б–Є–≤–љ–Њ–Љ —А–∞–Ј–Љ—Л–≤–µ —Б–Ї–ї–Њ–љ–Њ–≤ –і–Њ–ї–Є–љ—Л, —Б–ї–Њ–ґ–µ–љ–љ—Л—Е –∞–ї–µ–≤—А–Є—В–∞–Љ–Є. –Я–Њ–±–µ—А–µ–ґ—М–µ –Ј–∞–ї–Є–≤–∞ –Р—Е–Љ–∞—В–Њ–≤–∞ —В–µ—А—А–∞—Б–Є—А–Њ–≤–∞–љ–Њ вАУ –≤–і–Њ–ї—М –љ–µ–≥–Њ –њ–Њ –Њ–±–Њ–Є–Љ –±–µ—А–µ–≥–∞–Љ —Д—А–∞–≥–Љ–µ–љ—В–∞—А–љ–Њ –њ—А–Њ—Б–ї–µ–ґ–Є–≤–∞—О—В—Б—П –Љ–Њ—А—Б–Ї–Є–µ —В–µ—А—А–∞—Б—Л –≤—Л—Б–Њ—В–Њ–є 20 –Є 40вАУ50 –Љ, —З–∞—Б—В–Њ —Б–ї–Њ–ґ–µ–љ–љ—Л–µ —А—Л—Е–ї—Л–Љ –Љ–∞—В–µ—А–Є–∞–ї–Њ–Љ. –Ш—Б—Б–ї–µ–і–Њ–≤–∞–љ–љ–∞—П —А–µ–Ї–∞ —В–∞–Ї–ґ–µ –њ—А–Њ—А–µ–Ј–∞–µ—В —В–µ—А—А–∞—Б—Л 20 –Є 50 –Љ. –Я–µ—А–≤—Л–є —А–∞–Ј—А–µ–Ј –Љ–Њ—А—Б–Ї–Њ–є —В–µ—А—А–∞—Б—Л –≤—Л—Б–Њ—В–Њ–є 20 –Љ –Њ–±—А–∞–Ј–Њ–≤–∞–љ —Н—А–Њ–Ј–Є–µ–є –≤ 400 –Љ –Њ—В –Ї—А–∞—П –і–µ–ї—М—В—Л. –Ю–љ –Њ–Ї–∞–Ј–∞–ї—Б—П —Б–ї–Њ–ґ–µ–љ–љ—Л–Љ –∞–ї–µ–≤—А–Є—В–∞–Љ–Є —Б –њ—А–Њ—Б–ї–Њ—П–Љ–Є –≥–∞–ї—М–Ї–Є –Є —Б–Њ–ї–µ–є. –Т–≥–ї—Г–±—М –і–Њ–ї–Є–љ—Л –µ—С —Б–Ї–ї–Њ–љ—Л –њ—А–µ–і—Б—В–∞–≤–ї—П—О—В —Б–Њ–±–Њ–є –Њ—Б—Л–њ–љ—Л–µ —И–ї–µ–є—Д—Л —Б –Љ–Њ—А—Б–Ї–Є—Е —В–µ—А—А–∞—Б, –Ї–Њ—В–Њ—А—Л–µ –љ–∞ –Є–Ј–ї—Г—З–Є–љ–∞—Е —А–µ–Ї–Є –Њ–Ї–∞–Ј–∞–ї–Є—Б—М —А–∞–Ј–Љ—Л—В—Л–Љ–Є –Є –њ–Њ–і –љ–Є–Љ–Є –≤—Б–Ї—А—Л–ї–Є—Б—М —З—С—А–љ—Л–µ –≥–ї–Є–љ—Л, –Ї–Њ—В–Њ—А—Л–µ –Њ–Ї–Є—Б–ї—П—П—Б—М –≤ –њ—А–Њ—Ж–µ—Б—Б–µ –≤—Л—В–∞–Є–≤–∞–љ–Є—П, –њ—А–Є–Њ–±—А–µ—В–∞–ї–Є –Є–љ—В–µ–љ—Б–Є–≤–љ—Л–є —Б–µ—А—Л–є —Ж–≤–µ—В. –Ґ–∞–Ї–Њ–µ –Њ–±–љ–∞–ґ–µ–љ–Є–µ –Њ–±–љ–∞—А—Г–ґ–µ–љ–Њ –≤ 890 –Љ –Њ—В –Ї—А–∞—П –і–µ–ї—М—В—Л. –Т–Є–і–Є–Љ–∞—П –Љ–Њ—Й–љ–Њ—Б—В—М –≥–ї–Є–љ —Б–Њ—Б—В–∞–≤–ї—П–ї–∞ –Њ—В 4 –і–Њ 7 –Љ. –Э–∞ –≥–ї–Є–љ—Л —Б–≤–µ—А—Е—Г —Б –љ–µ—Б–Њ–≥–ї–∞—Б–Є–µ–Љ –ї–Њ–ґ–∞—В—Б—П –њ–µ—Б–Ї–Є –Є –∞–ї–µ–≤—А–Є—В—Л —Б –≥—А–∞–≤–Є–µ–Љ –Є –≥–∞–ї—М–Ї–Њ–є, –Ї–Њ—В–Њ—А—Л–µ –Є –Њ–±—А–∞–Ј—Г—О—В —В–µ–ї–Њ 50-–Љ–µ—В—А–Њ–≤–Њ–є —В–µ—А—А–∞—Б—Л. –Т —Н—В–Њ–є —В–Њ–ї—Й–µ –Њ–±–љ–∞—А—Г–ґ–Є–≤–∞–µ—В—Б—П –њ–ї–∞–≤–љ–Є–Ї –і—А–µ–≤–µ—Б–Є–љ—Л, –Ї–Њ—Б—В–Є –Љ–Њ—А—Б–Ї–Є—Е –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е, –Ї–Њ—В–Њ—А—Л–µ –Ј–∞–Ї–Њ–љ–Њ–Љ–µ—А–љ–Њ –Њ–Ї–∞–Ј—Л–≤–∞—О—В—Б—П –љ–∞ –і–љ–µ –і–Њ–ї–Є–љ—Л. –Ґ–∞–Ї–Є–Љ –Њ–±—А–∞–Ј–Њ–Љ, –≤—Б–Ї—А—Л—В—Л 3 –њ–∞—З–Ї–Є –Љ–Њ—А—Б–Ї–Є—Е –Њ—В–ї–Њ–ґ–µ–љ–Є–є, —Б–≤–Є–і–µ—В–µ–ї—М—Б—В–≤—Г—О—Й–Є–µ –Њ –њ—А–µ—А—Л–≤–Є—Б—В–Њ–Љ –Њ—Б–∞–і–Ї–Њ–љ–∞–Ї–Њ–њ–ї–µ–љ–Є–Є –≤ –Љ–Њ—А—Б–Ї–Є—Е —Г—Б–ї–Њ–≤–Є—П—Е –≤ –њ–Њ–Ј–і–љ–µ–Љ –љ–µ–Њ–њ–ї–µ–є—Б—В–Њ—Ж–µ–љ–µ.

–†–Є—Б. 1 –£—Б—В—М–µ –±–µ–Ј—Л–Љ—П–љ–љ–Њ–є —А–µ–Ї–Є, –≤–њ–∞–і–∞—О—Й–µ–є –≤ –Ј–∞–ї–Є–≤ –Р—Е–Љ–∞—В–Њ–≤–∞ —Б –Ј–∞–њ–∞–і–љ–Њ–≥–Њ –±–µ—А–µ–≥–∞. –Э–∞ –њ–µ—А–µ–і–љ–µ–Љ –њ–ї–∞–љ–µ –і–µ–ї—М—В–∞ —А–µ–Ї–Є, –љ–∞ –њ—А–∞–≤–Њ–Љ —Б–Ї–ї–Њ–љ–µ –і–Њ–ї–Є–љ—Л вАУ 20-–Љ–µ—В—А–Њ–≤–∞—П, –љ–∞ –ї–µ–≤–Њ–Љ 40-50 вАУ –Љ–µ—В—А–Њ–≤–∞—П –Љ–Њ—А—Б–Ї–Є–µ —В–µ—А—А–∞—Б—Л.

–†–Є—Б. 2 –Ф–Њ–ї–Є–љ–∞ –±–µ–Ј—Л–Љ—П–љ–љ–Њ–є —А–µ–Ї–Є, –≤–њ–∞–і–∞—О—Й–µ–є –≤ –Ј–∞–ї–Є–≤ –Р—Е–Љ–∞—В–Њ–≤–∞ —Б –Ј–∞–њ–∞–і–љ–Њ–≥–Њ –±–µ—А–µ–≥–∞. –Т–Є–і —Б–≤–µ—А—Е—Г вАУ –≤–љ–Є–Ј. –Т —Б–µ—А–µ–і–Є–љ–µ –њ–Њ–і–Љ—Л–≤–∞–µ–Љ–Њ–µ —А–µ–Ї–Њ–є –Њ–±–љ–∞–ґ–µ–љ–Є–µ —З—С—А–љ—Л—Е вАУ —Б–µ—А—Л—Е –≥–ї–Є–љ, —Г–≤–µ–ї–Є—З–Є–≤–∞—О—Й–Є—Е –≤–Є–і–Є–Љ—Г—О –Љ–Њ—Й–љ–Њ—Б—В—М —Б 4 –і–Њ 7 –Љ –≤–≤–µ—А—Е –њ–Њ —В–µ—З–µ–љ–Є—О —А–µ–Ї–Є. –Т–Є–і–µ–љ –љ–µ—А–Њ–≤–љ—Л–є –Ї–Њ–љ—В–∞–Ї—В –≥–ї–Є–љ –Є –љ–∞–ї–µ–≥–∞—О—Й–Є—Е –љ–∞ –љ–Є—Е –≥—А–∞–≤–Є–є–љ–Њ-–њ–µ—Б—З–∞–љ–Њ-–≥–∞–ї–µ—З–љ—Л—Е –Њ—В–ї–Њ–ґ–µ–љ–Є–є.
–Ґ–Њ—З–Ї–∞ –љ–∞–±–ї—О–і–µ–љ–Є—П (—В. –љ.) 3298.
09 –∞–≤–≥—Г—Б—В–∞ 2020 –≥.
–Ч–∞–њ–∞–і–љ—Л–є –±–µ—А–µ–≥ –Ј–∞–ї–Є–≤–∞ –Р—Е–Љ–∞—В–Њ–≤–∞, —Г—Б—В—М–µ —А—Г—З—М—П –≤ –≤–Є–і–µ –ї–Њ–њ–∞—Б—В–љ–Њ–є –і–µ–ї—М—В—Л –≤—Л–і–≤–Є–ґ–µ–љ–Є—П. –Ф–Њ–ї–Є–љ–∞ —А—Г—З—М—П вАУ —А–µ–Ї–Є —Г—Е–Њ–і–Є—В –≤ –≥–Њ—А—Л. (79¬∞04вАЩ04,6вАЭ —Б.—И 102¬∞44вАЩ43,0вАЭ –≤.–і.). –Ч–і–µ—Б—М –њ—А–Є–Љ–µ—А–љ–Њ –≤ 400 –Љ –Њ—В –Ї—А–∞—П –і–µ–ї—М—В—Л —А–µ–Ї–∞ –њ–Њ–і–Љ—Л–ї–∞ –њ—А–∞–≤—Л–є –±–µ—А–µ–≥ –Є –Њ–±–љ–∞–ґ–Є–ї–∞ —А–∞–Ј—А–µ–Ј –∞–ї–µ–≤—А–Є—В–Њ–≤ —Б –≤–µ—А—В–Є–Ї–∞–ї—М–љ–Њ–є —Б—В–µ–љ–Ї–Њ–є. –Т—Л—Б–Њ—В–∞ —Б—В–µ–љ–Ї–Є –і–Њ 8 –Љ. –Ю–±–љ–∞–ґ–µ–љ–Є–µ —Б–ї–Њ–ґ–µ–љ–Њ –≥–Њ—А–Є–Ј–Њ–љ—В–∞–ї—М–љ–Њ-—Б–ї–Њ–Є—Б—В—Л–Љ–Є —Б–µ—А–Њ-–ґ—С–ї—В—Л–Љ–Є –∞–ї–µ–≤—А–Є—В–∞–Љ–Є. –°–ї–Њ–Є—Б—В–Њ—Б—В—М –Њ–±–µ—Б–њ–µ—З–µ–љ–∞ —Ж–≤–µ—В–Њ–Љ вАУ –њ–µ—А–µ—Б–ї–∞–Є–≤–∞—О—В—Б—П –ґ—С–ї—В—Л–µ –Є —Б–µ—А—Л–µ —Б–ї–Њ–Є. –°–µ—А—Л–µ –і–Њ 2 —Б–Љ, –ґ–µ–ї—В—Л–µ –і–Њ 5 —Б–Љ. –†–∞–Ј—А–µ–Ј –љ–µ —А–∞—Б—З–Є—Й–µ–љ –њ–Њ–ї–љ–Њ—Б—В—М—О –Є–Ј-–∞ –љ–µ—Е–≤–∞—В–Ї–Є –≤—А–µ–Љ–µ–љ–Є. –Я–Њ—Н—В–Њ–Љ—Г –њ—А–Њ–Є–Ј–≤–µ–і–µ–љ–Њ –±–µ–≥–ї–Њ–µ –Њ–њ–Є—Б–∞–љ–Є–µ –Њ–±–љ–∞–ґ–µ–љ–Є—П. –Э–∞ –≤—Л—Б–Њ—В–µ 1,5вАУ1.8 –Љ –Њ—В –њ–Њ–і–Њ—И–≤—Л –Њ–±–љ–∞–ґ–µ–љ–Є—П (–і–љ–∞ –і–Њ–ї–Є–љ—Л) –≤ –∞–ї–µ–≤—А–Є—В–∞—Е –Ј–∞–ї–µ–≥–∞–µ—В –Ј–∞–Љ–µ—В–љ—Л–є –њ—А–Њ—Б–ї–Њ–є –Љ–µ–ї–Ї–Є—Е –≥–∞–ї–µ–Ї —В–Њ–ї—Й–Є–љ–Њ–є 25 —Б–Љ. –Ю—В–ї–Є—З–∞–µ—В—Б—П —В–µ–Љ, —З—В–Њ –≥–∞–ї—М–Ї–∞ –≤ –љ—С–Љ —Г–ї–Њ–ґ–µ–љ–∞ —Е–∞–Њ—В–Є—З–љ–Њ. –Э–∞ –≤—Л—Б–Њ—В–µ 6 –Љ –њ—А–Њ–Є–Ј–≤–µ–і–µ–љ–∞ —А–∞—Б—З–Є—Б—В–Ї–∞ –і–ї—П –Њ—В–±–Њ—А–∞ —Б—В–≤–Њ—А–Њ–Ї —А–∞–Ї–Њ–≤–Є–љ –Љ–Њ–ї–ї—О—Б–Ї–Њ–≤ –Є –њ–Њ—А–Њ–і—Л –і–ї—П –≠–Я–†-–∞–љ–∞–ї–Є–Ј–∞. –Ч–і–µ—Б—М —В–∞–Ї–ґ–µ –µ—Б—В—М –њ—А–Њ—Б–ї–Њ–є –Љ–µ–ї–Ї–Њ–є –≥–∞–ї—М–Ї–Є —В–Њ–ї—Й–Є–љ–Њ–є –Њ–Ї–Њ–ї–Њ 2 —Б–Љ, –Ї—А–Њ–Љ–µ —В–Њ–≥–Њ, –µ—Б—В—М –Њ—В–і–µ–ї—М–љ—Л–µ –≥–∞–ї—М–Ї–Є –≤ –њ–Њ—А–Њ–і–µ. –Т—Б—В—А–µ—З–∞—О—В—Б—П —В–Њ–љ–Ї–Њ—Б—В–µ–љ–љ—Л–µ —А–∞–Ї–Њ–≤–Є–љ—Л, –Ї–Њ—В–Њ—А—Л–µ –љ–µ–≤–Њ–Ј–Љ–Њ–ґ–љ–Њ –Њ—В–Њ–±—А–∞—В—М –±–µ–Ј —А–∞–Ј—А—Г—И–µ–љ–Є—П. –С–Њ–ї—М—И–Є–љ—Б—В–≤–Њ –Є–Ј –љ–Є—Е –Є–Љ–µ–µ—В –Њ—З–µ–љ—М –њ–µ—А–µ–ї–Є–≤—З–∞—В—Л–є –Є —А–∞–Ј–љ–Њ—Ж–≤–µ—В–љ—Л–є –њ–µ—А–ї–∞–Љ—Г—В—А–Њ–≤—Л–є —Б–ї–Њ–є. –†–∞–Ї–Њ–≤–Є–љ—Л –і–≤—Г—Б—В–≤–Њ—А—З–∞—В—Л–µ. –Я—А–Њ–Є–Ј–≤–µ–і—С–љ –Њ—В–±–Њ—А –Њ–±—А–∞–Ј—Ж–∞ —Н—В–Є—Е —А–∞–Ї–Њ–≤–Є–љ –Є –Њ—Б–Ї–Њ–ї–Ї–Њ–≤ –±–Њ–ї–µ–µ —В–Њ–ї—Б—В–Њ—Б—В–µ–љ–љ—Л—Е —А–∞–Ї–Њ–≤–Є–љ —Б –њ–Њ—А–Њ–і–Њ–є вАУ –Њ–±—А–∞–Ј–µ—Ж 3298/1. –Т—Л—И–µ —Б–ї–Њ—П –Њ—В–±–Њ—А–∞ –≤ 0,3 –Љ (–љ–∞ –≤—Л—Б–Њ—В–µ 6,5 –Љ) –≤ –∞–ї–µ–≤—А–Є—В–∞—Е –њ–Њ—П–≤–ї—П—О—В—Б—П –њ—А–Њ—Б–ї–Њ–Є —А–∞—Б—В–Є—В–µ–ї—М–љ–Њ—Б—В–Є —З—С—А–љ–Њ–≥–Њ —Ж–≤–µ—В–∞ –Є —Н—В–Є –њ—А–Њ—Б–ї–Њ–є–Ї–Є –≤ –њ–µ—А–≤—Л–µ –Љ–Є–ї–ї–Є–Љ–µ—В—А—Л-—Б–∞–љ—В–Є–Љ–µ—В—А—Л –≤—Б—В—А–µ—З–∞—О—В—Б—П –Є –≤—Л—И–µ вАУ –і–Њ –≤–µ—А—И–Є–љ—Л —Б—В–µ–љ–Ї–Є. –≠—В–Њ —В–Њ–љ–Ї–Є–µ —Б–њ—А–µ—Б–Њ–≤–∞–љ–љ—Л–µ, –≤–Є–і–Є–Љ–Њ –≤–Њ–ї–Њ–Ї–љ–∞ –≤–Њ–і–Њ—А–Њ—Б–ї–µ–є. –Ю—В–Њ–±—А–∞–љ –Њ–±—А–∞–Ј–µ—Ж —А–∞—Б—В–Є—В–µ–ї—М–љ–Њ—Б—В–Є —Б –њ–Њ—А–Њ–і–Њ–є 3298/2.
–Я—А–Њ–Є–Ј–≤–µ–і—С–љ –Њ—Б–Љ–Њ—В—А –Њ—В–ї–Њ–ґ–µ–љ–Є—П –≤—Л—И–µ –њ–Њ —В–µ—З–µ–љ–Є—О —А—Г—З—М—П –Є –Њ–±–љ–∞—А—Г–ґ–µ–љ —Б–ї–Њ–є —Б–Є–ї—М–љ–Њ –њ–∞—Е–љ—Г—Й–µ–є (—Б–µ—А–Њ–≤–Њ–і–Њ—А–Њ–і–Њ–Љ?) —З—С—А–љ–Њ–є –≥–ї–Є–љ—Л, –Ї–Њ—В–Њ—А–∞—П –Є –Ї—А–∞—Б–Є—В –≤–Њ–і—Г –≤ —А—Г—З—М–µ –≤ —В—С–Љ–љ–Њ-—Б–µ—А—Л–є —Ж–≤–µ—В. –Ґ–∞–Љ –ґ–µ –µ—Б—В—М –њ—А–Њ–і–Њ–ї–ґ–µ–љ–Є–µ —А–∞–Ј—А–µ–Ј–∞ –≤–≤–µ—А—Е. –Э–∞ –і–љ–µ –і–Њ–ї–Є–љ—Л –љ–∞–є–і–µ–љ—Л 2 –≤–ї–∞–ґ–љ—Л–µ –ї–µ–њ—С—И–Ї–Є —В–µ—Е —Б–∞–Љ—Л—Е —З—С—А–љ—Л—Е –≤–Њ–і–Њ—А–Њ—Б–ї–µ–є, –Ї–Њ—В–Њ—А—Л–µ –Љ–µ–ї–Ї–Є–Љ–Є –Ј–∞–ї–µ–ґ–∞–Љ–Є –Њ—В–Њ–±—А–∞–љ—Л –≤ –Њ–±—А. 3298/2. –Ы–µ–њ—С—И–Ї–Є –і–Њ 2 —Б–Љ —В–Њ–ї—Й–Є–љ–Њ–є, –Є –Њ–љ–Є –њ—А–∞–Ї—В–Є—З–µ—Б–Ї–Є —Б–Њ—Б—В–Њ—П—В –Є–Ј –Њ—А–≥–∞–љ–Є–Ї–Є. –Ю–±—А–∞–Ј–µ—Ж 3298/4 вАУ —З—С—А–љ—Л–µ –≤–Њ–і–Њ—А–Њ—Б–ї–Є –Є–Ј –≤–µ—А—Е–љ–µ–≥–Њ –Њ—В—А–µ–Ј–Ї–∞ –і–Њ–ї–Є–љ—Л. –Ю–Ї–∞—В—Л—И–Є –њ–Њ—А–Њ–і—Л —Б –≤–Њ–і–Њ—А–Њ—Б–ї—П–Љ–Є –љ–∞—Е–Њ–і—П—В—Б—П –љ–∞ –і–љ–Є—Й–µ –і–Њ–ї–Є–љ—Л. –Я—А–Є –Є—Е —А–∞–Ј–ї–∞–Љ—Л–≤–∞–љ–Є–Є —З—Г–≤—Б—В–≤—Г–µ—В—Б—П –Ј–∞–њ–∞—Е –є–Њ–і–∞.
–Э–∞ –і–љ–µ –і–Њ–ї–Є–љ—Л –љ–∞–є–і–µ–љ—Л –Љ–љ–Њ–≥–Њ—З–Є—Б–ї–µ–љ–љ—Л–µ —Б—В–≤–Њ—А–Ї–Є —А–∞–Ї–Њ–≤–Є–љ: Macoma, Hiatella, –Є –µ—Й—С –Ї–∞–Ї–∞—П-—В–Њ —Г–і–ї–Є–љ—С–љ–љ–∞—П —А–∞–Ї–Њ–≤–Є–љ–∞. –Ю–љ–Є –Њ—В–Њ–±—А–∞–љ—Л –≤ –Њ–±—А–∞–Ј–µ—Ж 3298/3.
–Э–∞ –і–љ–µ –і–Њ–ї–Є–љ—Л —В–∞–Ї–ґ–µ –Љ–љ–Њ–≥–Њ—З–Є—Б–ї–µ–љ–љ—Л–µ –Ї–Њ—Б—В–Є: 2 –њ–Њ–Ј–≤–Њ–љ–Ї–∞ –Ї–Є—В–∞, –љ–µ—Б–Ї–Њ–ї—М–Ї–Њ —А—С–±–µ—А, –њ–Њ-–≤–Є–і–Є–Љ–Њ–Љ—Г –Љ–Њ—А–ґ–∞ —Б—А–µ–і–љ–µ–є —Б–Њ—Е—А–∞–љ–љ–Њ—Б—В–Є. –Ю–љ–Є –Њ–Ї–∞—В–∞–љ—Л –њ—А–Є –њ–µ—А–µ–Љ–µ—Й–µ–љ–Є–Є –Є–Ј —А–∞–Ј—А–µ–Ј–∞ –Є –Є–Љ–µ—О—В —В–Є–њ–Є—З–љ—Л–є –±–µ–ї—Л–є —Ж–≤–µ—В. –Э–∞–є–і–µ–љ —В–∞–Ї–ґ–µ –Њ–і–Є–љ –Ї–ї—Л–Ї –Љ–Њ—А–ґ–∞ –Њ—В–ї–Є—З–љ–Њ–є —Б–Њ—Е—А–∞–љ–љ–Њ—Б—В–Є, —Б —Н–Љ–∞–ї—М—О, –Ї–Њ—А–Є—З–љ–µ–≤–∞—В–Њ–≥–Њ —Ж–≤–µ—В–∞ —Б —З–∞—Б—В—М—О —З–µ–ї—О—Б—В–Є.
13 –∞–≤–≥—Г—Б—В–∞ 2020 –≥.
–Я—А–Њ–і–Њ–ї–ґ–µ–љ–Є–µ –Њ–њ–Є—Б–∞–љ–Є—П –Њ–±–љ–∞–ґ–µ–љ–Є—П –љ–∞ –Ј–∞–њ–∞–і–љ–Њ–Љ –±–µ—А–µ–≥—Г –Ј–∞–ї–Є–≤–∞ –Р—Е–Љ–∞—В–Њ–≤–∞ (79¬∞04вАЩ11,3вАЭ —Б.—И 102¬∞43вАЩ33,0вАЭ –≤.–і.) —В–Њ—З–Ї–∞ –љ–∞—Е–Њ–і–Є—В—Б—П –≤ –≥–ї—Г–±–Є–љ–µ –і–Њ–ї–Є–љ—Л –љ–∞ —А–∞—Б—Б—В–Њ—П–љ–Є–Є 890 –Љ –Њ—В –Ї—А–∞—П –і–µ–ї—М—В—Л —А–µ–Ї–Є (–±–µ—А–µ–≥–∞ –Љ–Њ—А—П). –Т—Л—Б–Њ—В–∞ –њ–Њ–і–Њ—И–≤—Л –Њ–±–љ–∞–ґ–µ–љ–Є—П –њ–Њ —Н–Ї–ї–Є–Љ–µ—В—А–Є—А–Њ–≤–∞–љ–Є—О вАУ 25 –Љ, –≤—Л—Б–Њ—В–∞ –њ–Њ –±–∞—А–Њ–Љ–µ—В—А—Г 25 –Љ, –њ–Њ GPS - 23 –Љ. –Я–Њ–і–љ–Њ–ґ–Є–µ –Њ–±–љ–∞–ґ–µ–љ–Є—П I, –Њ–њ–Є—Б–∞–љ–љ–Њ–≥–Њ 9 –∞–≤–≥—Г—Б—В–∞, —А–∞—Б–њ–Њ–ї–Њ–ґ–µ–љ–Њ –љ–∞ –≤—Л—Б–Њ—В–µ 9 –Љ. –Ю–±–љ–∞–ґ–µ–љ–Є–µ II —Б –њ–Њ–і–Њ—И–≤–Њ–є –љ–∞ –≤—Л—Б–Њ—В–µ 25 –Љ —Е–∞—А–∞–Ї—В–µ—А–љ–Њ –Њ—В–ї–Є—З–љ–Њ–є –≤—Б–Ї—А—Л—В–Њ—Б—В—М—О —А—Г—З—М—С–Љ —Б—В–µ–љ–Ї–Є —З—С—А–љ—Л—Е –≥–ї–Є–љ, –Ї–Њ—В–Њ—А—Л–µ –њ–µ—А–µ–Ї—А—Л–≤–∞—О—В—Б—П –њ–µ—Б—З–∞–љ–Њ-–≥–∞–ї–µ—З–љ–Њ-–≥—А–∞–≤–Є–є–љ–Њ–є —В–Њ–ї—Й–µ–є.
–Т—Л—И–µ –њ–Њ —В–µ—З–µ–љ–Є—О –≥–ї–Є–љ—Л —Г–≤–µ–ї–Є—З–Є–≤–∞—О—В –≤–Є–і–Є–Љ—Г—О –Љ–Њ—Й–љ–Њ—Б—В—М —Б 4 –і–Њ 7 –Љ –Є –і–∞–ї–µ–µ –≤—Л–Ї–ї–Є–љ–Є–≤–∞—О—В—Б—П –≤–≤–µ—А—Е –њ–Њ –і–Њ–ї–Є–љ–µ, –Ј–∞–Ї—А—Л–≤–∞—П—Б—М –Њ—Б—Л–њ–љ—Л–Љ–Є —И–ї–µ–є—Д–∞–Љ–Є –њ–µ—Б—З–∞–љ–Њ-–≥–∞–ї–µ—З–љ–Њ–є —В–Њ–ї—Й–Є, —Б–ї–∞–≥–∞—О—Й–µ–є —В–µ—А—А–∞—Б—Г –≤—Л—Б–Њ—В–Њ–є 51 –Љ –≤ –±—А–Њ–≤–Ї–µ, 60 –Љ –≤ —В—Л–ї–Њ–≤–Њ–Љ —И–≤–µ. –Э–∞ –њ–Њ–≤–µ—А—Е–љ–Њ—Б—В–Є —В–µ—А—А–∞—Б—Л –Ј–∞–ї–µ–≥–∞—О—В —Е–Њ—А–Њ—И–Њ –Є –Њ—В–ї–Є—З–љ–Њ –Њ–Ї–∞—В–∞–љ–љ—Л–µ –≥–∞–ї—М–Ї–Є.
–І—С—А–љ—Л–µ –≥–ї–Є–љ—Л –≤—Б–Ї—А—Л—В—Л —А—Г—З—М—С–Љ –њ–Њ–і –ї–µ–≤—Л–Љ —Б–Ї–ї–Њ–љ–Њ–Љ –і–Њ–ї–Є–љ—Л –Є –Њ–±—А–∞–Ј—Г—О—В –≤—Л–њ—Г–Ї–ї—Г—О —Б—В–µ–љ–Ї—Г –≤—Л—Б–Њ—В–Њ–є 3,5вАУ4 –Љ. –Ю–љ–Є –њ—А–µ–і—Б—В–∞–≤–ї–µ–љ—Л –∞–ї–µ–≤—А–Є—В–Њ–≤–Њ–є –≥–ї–Є–љ–Њ–є —З—С—А–љ–Њ–≥–Њ —Ж–≤–µ—В–∞, –Ї–Њ—В–Њ—А–∞—П, –±—Л—Б—В—А–Њ –Њ–Ї–Є—Б–ї—П—П—Б—М –Љ–µ–љ—П–µ—В —Ж–≤–µ—В –љ–∞ —Б–µ—А—Л–є. –Т —Н—В–Њ–є —Б—В–µ–љ–Ї–µ –њ—А–Њ–Є—Б—Е–Њ–і–Є—В –∞–Ї—В–Є–≤–љ–Њ–µ –Њ–±–≤–∞–ї–Є–≤–∞–љ–Є–µ –Њ–±–љ–∞–ґ–µ–љ–Є—П –њ–ї–∞—Б—В–Є–љ–Ї–∞–Љ–Є –≤ —А–µ–Ј—Г–ї—М—В–∞—В–µ –љ–∞–≥—А–µ–≤–∞–љ–Є—П, –Њ—Б–Њ–±–µ–љ–љ–Њ –њ–Њ–і –ї—Г—З–∞–Љ–Є —Б–Њ–ї–љ—Ж–∞. –≠—В–Њ –њ—А–Њ–Є—Б—Е–Њ–і–Є—В –њ–Њ—В–Њ–Љ—Г, —З—В–Њ –≥–ї–Є–љ—Л –љ–∞—Б—Л—Й–µ–љ—Л –ї—М–і–Њ–Љ (–і–Њ 20вАУ30%), –њ—А–Є—З—С–Љ –ї–µ–і—П–љ—Л–Љ–Є –≤–µ—А—В–Є–Ї–∞–ї—М–љ—Л–Љ–Є –ґ–Є–ї–Ї–∞–Љ–Є –≤ –≤–Є–і–µ –Љ–љ–Њ–≥–Њ–Ј—Г–±—Ж–µ–≤, –Њ–±—А–∞—Й—С–љ–љ—Л—Е –Њ—Б—В—А–Є—П–Љ–Є –≤–≤–µ—А—Е –Є —Б –Њ–±—Й–Є–Љ –ї—М–і–Є—Б—В—Л–Љ –Њ—Б–љ–Њ–≤–∞–љ–Є–µ–Љ. –Т—Л—Б–Њ—В–∞ —В–∞–Ї–Њ–≥–Њ –Њ—В–і–µ–ї—М–љ–Њ–≥–Њ —В–µ–ї–∞ 0,7 –Љ, —И–Є—А–Є–љ–∞ 0,6 –Љ.
–Р–ї–µ–≤—А–Є—В–Њ–≤—Л–µ –≥–ї–Є–љ—Л –Њ—В–ї–Є—З–љ–Њ —Б–ї–Њ–Є—Б—В—Л, —В–Њ–ї—Й–Є–љ–∞ —Б–ї–Њ–є–Ї–Њ–≤ –Њ—В –њ–µ—А–≤—Л—Е –Љ–Є–ї–ї–Є–Љ–µ—В—А–Њ–≤ –і–Њ 1 —Б–Љ. –Э–Њ –≤ –љ–Є—Е –µ—Б—В—М –љ–µ–Ї–Њ—В–Њ—А–∞—П –љ–µ–Њ–і–љ–Њ—А–Њ–і–љ–Њ—Б—В—М, —З—В–Њ –њ—А–Є–і–∞—С—В –≥–ї–Є–љ–∞–Љ –≤–Њ–Ј–і—Г—И–љ–Њ—Б—В—М –њ—А–Є —Б–Љ–Є–љ–∞–љ–Є–Є. –Т–µ—А–Њ—П—В–љ–µ–µ –≤—Б–µ–≥–Њ —Н—В–Њ –њ—А–Њ–Є—Б—Е–Њ–і–Є—В –Є–Ј-–Ј–∞ –љ–∞–ї–Є—З–Є—П –≤ –≥–ї–Є–љ–∞—Е –Њ—Б—В–∞—В–Ї–Њ–≤ —А–∞—Б—В–Є—В–µ–ї—М–љ–Њ—Б—В–Є, –∞ –Є–Љ–µ–љ–љ–Њ –≤–Њ–ї–Њ–Ї–Њ–љ –≤–Њ–і–Њ—А–Њ—Б–ї–µ–є, –Њ—В–њ–µ—З–∞—В–Ї–Є –Є —Б–∞–Љ–Є —В–Њ–љ–Ї–Є–µ –≤–Њ–ї–Њ–Ї–љ–∞ –Ї–Њ—В–Њ—А—Л—Е –Є–љ–Њ–≥–і–∞ –Њ–±–љ–∞—А—Г–ґ–Є–≤–∞—О—В—Б—П –њ—А–Є —А–∞–Ј–і–µ–ї–µ–љ–Є–Є —Б–ї–Њ–є–Ї–Њ–≤. –†—Г—З–µ–є –љ–∞ —Г—А–µ–Ј–µ –≤–Њ–і—Л –Є –≤—Л—И–µ –љ–∞ 0,5 –Љ –≤—Б–Ї—А—Л–ї —А–∞–Ї—Г—И–љ—П–Ї–Њ–≤—Г—О –±–∞–љ–Ї—Г, –њ–Њ—Б—В–Њ—П–љ–љ–Њ –њ—А–µ–њ–∞—А–Є—А—Г—П –Є –≤—Л–Љ—Л–≤–∞—П —А–∞–Ї–Њ–≤–Є–љ—Л –і–≤—Г—Б—В–≤–Њ—А—З–∞—В—Л—Е –Љ–Њ–ї–ї—О—Б–Ї–Њ–≤. –†–∞–Ї–Њ–≤–Є–љ—Л —А–∞—Б–њ–Њ–ї–Њ–ґ–µ–љ—Л –≤ –±–µ—Б–њ–Њ—А—П–і–Њ—З–љ–Њ–Љ –њ–Њ–ї–Њ–ґ–µ–љ–Є–Є, –Ї–Њ—В–Њ—А—Л–µ –≤–µ—А—В–Є–Ї–∞–ї—М–љ–Њ, –Ї–Њ—В–Њ—А—Л–µ –≥–Њ—А–Є–Ј–Њ–љ—В–∞–ї—М–љ–Њ –Є–ї–Є –љ–∞–Ї–ї–Њ–љ–љ–Њ –Ј–∞–ї–µ–≥–∞—О—В –њ–Њ –Њ—В–љ–Њ—И–µ–љ–Є—О –Ї —Б–ї–Њ–Є—Б—В–Њ—Б—В–Є. –Ш–ї–Є —Б–∞–Љ–Є —Б–ї–Њ–Є –Є–Љ–µ—О—В –љ–∞—А—Г—И–µ–љ–Є–µ –≥–Њ—А–Є–Ј–Њ–љ—В–∞–ї—М–љ–Њ–≥–Њ –Ј–∞–ї–µ–≥–∞–љ–Є—П, —З–µ–≥–Њ –≤ —Н—В–Њ–є —З–∞—Б—В–Є –Њ–±–љ–∞–ґ–µ–љ–Є—П –љ–µ –Њ–њ—А–µ–і–µ–ї–Є—В—М. –І–∞—Б—В–Њ —А–∞–Ї–Њ–≤–Є–љ—Л –њ—А–Є—Б–ї–Њ–љ–µ–љ—Л –і—А—Г–≥ –Ї –і—А—Г–≥—Г. –Ю–љ–Є –Ј–∞–њ–Њ–ї–љ–µ–љ—Л –ї—М–і–Є—Б—В–Њ–є –њ–Њ—А–Њ–і–Њ–є –Є–ї–Є –ї—М–і–Њ–Љ, –Њ—В—З–µ–≥–Њ –Ї–∞–ґ—Г—Й–Є–µ—Б—П –њ—А–Њ—З–љ—Л–Љ–Є, –±—Л—Б—В—А–Њ —А–∞–Ј—А—Г—И–∞—О—В—Б—П –њ—А–Є –≤—Л—В–∞–Є–≤–∞–љ–Є–Є –ї—М–і–∞ –Є —Б –њ–Њ—В–µ—А–µ–є –Ї–Њ–љ—В–∞–Ї—В–∞ —Б –љ–∞—Е–Њ–і—П—Й–µ–є—Б—П –≤–љ—Г—В—А–Є —Б—В–≤–Њ—А–Њ–Ї –њ–Њ—А–Њ–і–Њ–є. –Х—Б—В—М —А–∞–Ї–Њ–≤–Є–љ—Л –њ–Њ–ї–љ—Л–µ –Є–Ј –і–≤—Г—Е —Б—В–≤–Њ—А–Њ–Ї. –Э–µ—Б–Њ–Љ–љ–µ–љ–љ–Њ, –Љ–µ—А–Ј–ї–Њ—В–љ—Л–µ –њ—А–Њ—Ж–µ—Б—Б—Л –Њ—В–≤–µ—В—Б—В–≤–µ–љ–љ—Л –Ј–∞ —В—А–µ—Й–Є–љ–Њ–≤–∞—В–Њ—Б—В—М –Є —А–∞–Ј—А—Г—И–µ–љ–Є–µ —А–∞–Ї–Њ–≤–Є–љ. –†–∞–Ї–Њ–≤–Є–љ—Л –і–≤—Г—Б—В–≤–Њ—А—З–∞—В—Л—Е –Љ–Њ–ї–ї—О—Б–Ї–Њ–≤ –≤ –≥–ї–Є–љ–∞—Е —В–Њ–љ–Ї–Њ—Б—В–µ–љ–љ—Л–µ, –≤ –Њ—В–ї–Є—З–Є–µ –Њ—В —В–Њ–ї—Б—В–Њ—Б—В–µ–љ–љ—Л—Е, –њ–Њ–≤—Б—О–і—Г —А–∞—Б–Ї–Є–і–∞–љ–љ—Л—Е –њ–Њ –і–љ—Г –і–Њ–ї–Є–љ—Л. –Ш–Ј —Н—В–Њ–є —З–∞—Б—В–Є –Њ–±–љ–∞–ґ–µ–љ–Є—П –≤ –Є–љ—В–µ—А–≤–∞–ї–µ –Њ—В —Г—А–µ–Ј–∞ –≤–Њ–і—Л –і–Њ 0,5 –Љ –≤—Л—И–µ –љ–µ–≥–Њ –Њ—В–Њ–±—А–∞–љ—Л –Њ–±—А–∞–Ј—Ж—Л: 3298/5 вАУ –њ–Њ—А–Њ–і–∞ -–∞–ї–µ–≤—А–Є—В–Њ–≤–∞—П –≥–ї–Є–љ–∞ —Б–Њ —Б—В–≤–Њ—А–Ї–∞–Љ–Є —А–∞–Ї–Њ–≤–Є–љ –Љ–Њ–ї–ї—О—Б–Ї–Њ–≤ –і–ї—П –≠–Я–†-–∞–љ–∞–ї–Є–Ј–∞ –≤–Њ–Ј—А–∞—Б—В–∞ –≤ –њ-—Н—В. –њ–∞–Ї–µ—В. 3298/6 вАУ –≥–ї–Є–љ—Л –і–ї—П –Љ–Є–Ї—А–Њ—Д–∞—Г–љ–Є—Б—В–Є—З–µ—Б–Ї–Њ–≥–Њ –Є –і–Є–∞—В–Њ–Љ–Њ–≤–Њ–≥–Њ –∞–љ–∞–ї–Є–Ј–∞ –≤ –њ-—Н—В. –њ–∞–Ї–µ—В, 3298/7 вАУ —А–∞–Ї–Њ–≤–Є–љ—Л –і–ї—П –Њ–њ—А–µ–і–µ–ї–µ–љ–Є—П –≤–Є–і–Њ–≤ –Љ–Њ–ї–ї—О—Б–Ї–Њ–≤ –≤ 2 –њ-—Н—В. –њ–∞–Ї–µ—В–∞.
–Я–Њ—Б–ї–µ–і–љ—П—П –њ—А–Њ–±–∞ —Б–Њ–±–Є—А–∞–ї–∞—Б—М —В–∞–Ї, —З—В–Њ–±—Л –≤ —А–∞–Ї–Њ–≤–Є–љ–∞—Е –Њ—Б—В–∞–≤–∞–ї–∞—Б—М —Б–Ї—А–µ–њ–ї—П—О—Й–∞—П –Є—Е –≥–ї–Є–љ–∞. –Э–Њ –≤ –њ–∞–Ї–µ—В–µ –њ—А–Њ–Є–Ј–Њ—И–ї–Њ –≤—Л—В–∞–Є–≤–∞–љ–Є–µ –ї—М–і–∞ –Є —А–∞–Ї–Њ–≤–Є–љ—Л —А–∞–Ј—А—Г—И–Є–ї–Є—Б—М. –°—Г–і—П –њ–Њ —Н—В–Є–Љ –њ—А–Њ–±–∞–Љ –≤–ї–∞–ґ–љ–Њ—Б—В—М –≥–ї–Є–љ –≤—Л—Б–Њ–Ї–∞ вАУ –љ–µ –Љ–µ–љ–µ–µ 20%. –Т—Б–µ –њ—А–Њ–±—Л –њ—А–µ–≤—А–∞—В–Є–ї–Є—Б—М –≤ —В–µ—Б—В–Њ–њ–Њ–і–Њ–±–љ—Г—О –њ–Њ–ї—Г–ґ–Є–і–Ї—Г—О –Љ–∞—Б—Б—Г. –Э–Њ –≤–Њ –≤—А–µ–Љ—П –Њ—В–Љ—Л–≤–Ї–Є —А–∞–Ї–Њ–≤–Є–љ —Г–ґ–µ –≤ –љ–Њ—П–±—А–µ 2020 –≥. —Г–і–∞–ї–Њ—Б—М –њ–Њ–ї—Г—З–Є—В—М 3 –њ–Њ–ї–љ—Л–µ –і–≤—Г—Б—В–≤–Њ—А–Ї–Є —Б –Ј–∞–њ–Њ–ї–љ—П—О—Й–Є–Љ –Є—Е –≥–ї–Є–љ–Є—Б—В—Л–Љ –∞–ї–µ–≤—А–Є—В–Њ–Љ, –љ–µ—Б–Ї–Њ–ї—М–Ї–Њ –Њ–і–Є–љ–Њ—З–љ—Л—Е —Б—В–≤–Њ—А–Њ–Ї –Є –Њ—Б–Ї–Њ–ї–Ї–Є —А–∞–Ј–љ–Њ–≥–Њ —А–∞–Ј–Љ–µ—А–∞. –Т—Б—П –њ—А–Њ–±–∞ –Њ—В–і–∞–љ–∞ –њ–∞–ї–µ–Њ–љ—В–Њ–ї–Њ–≥—Г –Р. –Т. –Ъ—А—Л–ї–Њ–≤—Г –і–ї—П –Њ–њ—А–µ–і–µ–ї–µ–љ–Є—П –≤–Є–і–Њ–≤–Њ–≥–Њ —Б–Њ—Б—В–∞–≤–∞ –і–≤—Г—Б—В–≤–Њ—А—З–∞—В—Л—Е –Љ–Њ–ї–ї—О—Б–Ї–Њ–≤. –Ш–Ј –љ–µ—Б–Ї–Њ–ї—М–Ї–Є—Е –Њ–і–Є–љ–Њ—З–љ—Л—Е —Б—В–≤–Њ—А–Њ–Ї —Г–і–∞–ї–Њ—Б—М –Є–Ј—К—П—В—М –Ј–∞–њ–Њ–ї–љ—П—О—Й–Є–є –Є—Е –≥–ї–Є–љ–Є—Б—В—Л–є –∞–ї–µ–≤—А–Є—В –≤ –Њ—В–і–µ–ї—М–љ—Л–є –њ–∞–Ї–µ—В 3298/7 –і–ї—П –і—А—Г–≥–Є—Е –≤–Є–і–Њ–≤ –∞–љ–∞–ї–Є–Ј–∞.
–Т—Л—И–µ –њ–Њ –і–Њ–ї–Є–љ–µ –љ–∞ 50-100 –Љ –Є –њ—А–Є –≤—Л—Б–Њ—В–µ —Г—А–µ–Ј–∞ –≤–Њ–і—Л 26,7 –Љ –љ–∞–і —Г—А–Њ–≤–љ–µ–Љ –Љ–Њ—А—П –≥–ї–Є–љ—Л –≤—Б–Ї—А—Л–≤–∞—О—В—Б—П –љ–∞ –±–Њ–ї—М—И—Г—О –Љ–Њ—Й–љ–Њ—Б—В—М вАУ –і–Њ 7 –Љ. –Ш–Ј —Н—В–Њ–є —Б—В–µ–љ–Ї–Є —Г–ґ–µ —Б–µ—А—Л—Е –Њ–Ї–Є—Б–ї–Є–≤—И–Є—Е—Б—П –≥–ї–Є–љ –Њ—В–Њ–±—А–∞–љ—Л –њ—А–Њ–±—Л —Б –≥–Њ—А–Є–Ј–Њ–љ—В–∞ 29,2 –Љ –љ–∞–і —Г.–Љ. (2,5 –Љ –љ–∞–і —Г—А–µ–Ј–Њ–Љ –≤–Њ–і—Л –≤ —А—Г—З—М–µ) вАУ –Њ–±—А–∞–Ј–µ—Ж 3298/8, –Є —Б –≤—Л—Б–Њ—В—Л 33,7 –Љ, –Є–Ј –Ї—А–Њ–≤–ї–Є –≥–ї–Є–љ вАУ –Њ–±—А. 3298/9. –Ъ—А–Њ–≤–ї—П –≥–ї–Є–љ –љ–µ—А–Њ–≤–љ–∞—П, –љ–Њ –±–µ–Ј –≤—Б—П–Ї–Є—Е –Ј–∞—В—С–Ї–Њ–≤ –Є –Ї–∞—А–Љ–∞–љ–Њ–≤.
–Т—Л—И–µ –Ј–∞–ї–µ–≥–∞–µ—В —В–Њ–ї—Й–∞ –њ–µ—А–µ—Б–ї–∞–Є–≤–∞–љ–Є—П –≥—А–∞–≤–Є—П –Є –њ–ї–∞—Б—В–Њ–≤ —Б–µ—А–Њ–≥–Њ –њ–µ—Б–Ї–∞, –ґ–µ–ї—В–Њ–≤–∞—В–Њ-—Б–µ—А–Њ–≥–Њ –∞–ї–µ–≤—А–Є—В–∞ –Є —А–∞—Б—В–Є—В–µ–ї—М–љ–Њ—Б—В–Є (–≤–µ—А–Њ—П—В–љ–Њ, –≤–Њ–і–Њ—А–Њ—Б–ї–µ–є —З—С—А–љ–Њ–≥–Њ —Ж–≤–µ—В–∞. –≠—В–Є –њ–∞—З–Ї–Є —Б–Њ—Б—В–Њ—П—В –Є–Ј —А–∞—Б—В–Є—В–µ–ї—М–љ–Њ—Б—В–Є —В–Њ–ї—Й–Є–љ–Њ–є –њ—А–Њ—Б–ї–Њ–µ–≤ –і–Њ 2вАУ3 —Б–Љ, —Б–ї–Њ–є–Ї–Њ–≤ –∞–ї–µ–≤—А–Є—В–Њ–≤ –і–Њ 10 —Б–Љ –Є –њ–µ—Б–Ї–Њ–≤ —Б –≥—А–∞–≤–Є–µ–Љ —В–Њ–ї—Й–Є–љ–Њ–є –≤ –њ–µ—А–≤—Л–µ —Б–∞–љ—В–Є–Љ–µ—В—А—Л. –Я–∞—З–Ї–∞ –≤ —Ж–µ–ї–Њ–Љ –Є–Љ–µ–µ—В —В–Њ–ї—Й–Є–љ—Г 15-20 —Б–Љ. –Ґ–∞–Ї–Є—Е –њ–∞—З–µ–Ї –љ–∞–і –≥–ї–Є–љ–∞–Љ–Є —Б—А–µ–і–Є –≥—А–∞–≤–Є–є–љ–Є–Ї–Њ–≤ –љ–∞—Б—З–Є—В—Л–≤–∞–µ—В—Б—П 3, –≤ –љ–µ–Ї–Њ—В–Њ—А—Л—Е –Љ–µ—Б—В–∞—Е –љ–∞ —Б–Ї–ї–Њ–љ–µ вАУ 4. –Ґ–∞–Ї –Ї–∞–Ї –Њ—Б—Л–њ–љ–Њ–є —Б–Ї–ї–Њ–љ –Є–Љ–µ–µ—В –Ї—А—Г—В–Є–Ј–љ—Г 30-35¬∞ –Є —Б–ї–Њ–ґ–µ–љ –≥–∞–ї–µ—З–љ–Њ-–≥—А–∞–≤–Є–є–љ—Л–Љ –Љ–∞—В–µ—А–Є–∞–ї–Њ–Љ –≤—Б–µ —Б–ї–Њ–Є –Њ—Б–∞–і–Ї–Њ–≤ —Б–Љ–µ—Й–µ–љ—Л –≤–љ–Є–Ј –њ–Њ —Б–Ї–ї–Њ–љ—Г, –∞ —Н—В–Є –њ–∞—З–Ї–Є –њ–µ—А–µ—А–µ—Б–ї–∞–Є–≤–∞–љ–Є—П –≤–Њ–і–Њ—А–Њ—Б–ї–µ–є, –њ–µ—Б–Ї–Њ–≤ –Є –∞–ї–µ–≤—А–Є—В–Њ–≤ –±–µ–Ј —А–∞–Ј—А—Л–≤–∞ —Б–њ–ї–Њ—И–љ–Њ—Б—В–Є, –љ–Њ –≤—Б–Ї—А—Л–≤–∞—О—В—Б—П –Њ–љ–Є —Б–ї–µ–≥–Ї–∞ –љ–∞ —А–∞–Ј–љ–Њ–є –≤—Л—Б–Њ—В–µ, –Є–Ј–Љ–µ–љ—П—П –µ—С –≤ –њ—А–µ–і–µ–ї–∞—Е 1-2 –Љ. –Ъ–Њ–љ—В–∞–Ї—В –≥—А–∞–≤–Є–є–љ–Є–Ї–Њ–≤ —Б –њ–Њ–і—Б—В–Є–ї–∞—О—Й–Є–Љ–Є –≥–ї–Є–љ–∞–Љ–Є –љ–µ—А–Њ–≤–љ—Л–є. –У—А–∞–≤–Є–є–љ–Є–Ї–Є –њ–Њ—Б—В–Њ—П–љ–љ–Њ –љ–∞–і–≤–Є–≥–∞—О—В—Б—П –љ–∞ –±—А–Њ–≤–Ї—Г —Б—В–µ–љ–Ї–Є, —Б–ї–Њ–ґ–µ–љ–љ—Г—О –≥–ї–Є–љ–∞–Љ–Є, –Є –Њ—Б—Л–њ–∞—О—В—Б—П –і–∞–ї–µ–µ –і–Њ –і–љ–∞ —А—Г—З—М—П. –Ш–Ј –Ј–Њ–љ—Л –Ї–Њ–љ—В–∞–Ї—В–∞, –љ–∞ 0,8 –Љ –≤—Л—И–µ –≥–ї–Є–љ (34.5 –Љ –љ–∞–і —Г—А. –Љ–Њ—А—П) –Є–Ј –≥—А–∞–≤–Є–є–љ–Є–Ї–Њ–≤ –≤–Ј—П—В –Њ–±—А–∞–Ј–µ—Ж 3298/10. –Э–∞ –≤—Л—Б–Њ—В–µ 2 –Љ –≤—Л—И–µ –Ї–Њ–љ—В–∞–Ї—В–∞ —Б –≥–ї–Є–љ–∞–Љ–Є (35,7 –Љ –љ–∞–і —Г—А. –Љ–Њ—А—П) –Њ—В–Њ–±—А–∞–љ—Л –Њ–±—А–∞–Ј—Ж—Л –Є–Ј –њ–∞—З–Ї–Є –њ–µ—А–µ—Б–ї–∞–Є–≤–∞–љ–Є—П –њ–µ—Б–Ї–Њ–≤, –∞–ї–µ–≤—А–Є—В–Њ–≤, –≤–Њ–і–Њ—А–Њ—Б–ї–µ–є: 3298/11 вАУ –∞–ї–µ–≤—А–Є—В —Б –≥—А–∞–≤–Є–µ–Љ –≤ –њ–∞–Ї–µ—В, 3298/11 –Ю–°–Ы- –∞–ї–µ–≤—А–Є—В —Б –≥—А–∞–≤–Є–µ–Љ –≤ —В—А—Г–±–Ї—Г –і–ї—П –њ—А–Њ–≤–µ–і–µ–љ–Є—П –Ю–°–Ы-–∞–љ–∞–ї–Є–Ј–∞ –≤–Њ–Ј—А–∞—Б—В–∞, 3298/12 –і—А–µ–≤–µ—Б–Є–љ—Л –Є–Ј –њ–∞—З–Ї–Є –њ–µ—А–µ—Б–ї–∞–Є–≤–∞–љ–Є—П –і–ї—П —А–∞–і–Є–Њ—Г–≥–ї–µ—А–Њ–і–љ–Њ–≥–Њ –і–∞—В–Є—А–Њ–≤–∞–љ–Є—П –Ю–±—А–∞–Ј–µ—Ж 3298/13 вАУ –Ї–Њ—Б—В—М –Љ–Њ—А—Б–Ї–Њ–≥–Њ –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–µ–≥–Њ (—А–µ–±—А–Њ?) –Є–Ј –Њ—Б—Л–њ–Є –љ–∞ —Н—В–Њ–Љ –ґ–µ –≤—Л—Б–Њ—В–љ–Њ–Љ —Г—А–Њ–≤–љ–µ. –Ф—А–µ–≤–µ—Б–љ—Л–є —Б—В–≤–Њ–ї –і–Є–∞–Љ–µ—В—А–Њ–Љ 6 —Б–Љ —В–Њ—А—З–∞–ї –Є–Ј –њ–∞—З–Ї–Є –њ–µ—А–µ—Б–ї–∞–Є–≤–∞–љ–Є—П –≤–Њ–і–Њ—А–Њ—Б–ї–µ–є, –∞–ї–µ–≤—А–Є—В–Њ–≤, –њ–µ—Б–Ї–Њ–≤ –љ–∞ 1,2 –Љ. –Я—А–Њ–Є–Ј–≤–µ–і–µ–љ–Њ —Г–≥–ї—Г–±–ї–µ–љ–Є–µ –і–Њ –Љ–µ—А–Ј–ї–Њ—В—Л –≤ —Б—В–µ–љ–Ї–µ (–љ–∞, 0,4 –Љ) –Є –Є–Ј–≤–ї–µ—З–µ–љ–∞ –њ–Њ–≥—А–µ–±—С–љ–љ–∞—П —З–∞—Б—В—М —Б—В–≤–Њ–ї–∞. –Ф—А–µ–≤–µ—Б–Є–љ–∞ —Б—В–∞—А–∞—П –Є –Њ—В—Б–ї–∞–Є–≤–∞–µ—В—Б—П –њ–Њ –≥–Њ–і–Є—З–љ—Л–Љ –Ї–Њ–ї—М—Ж–∞–Љ –Ї–∞–Ї –Ї–Њ—А–∞. –Я–Њ—Б–ї–µ–і—Г—О—Й–∞—П —А–∞—Б–њ–Є–ї–Њ–≤–Ї–∞ –і—А–µ–≤–µ—Б–Є–љ—Л –њ–Њ–Ї–∞–Ј–∞–ї–∞, —З—В–Њ –і—А–µ–≤–µ—Б–Є–љ–∞ –љ–∞—Е–Њ–і–Є—В—Б—П –≤ —Б—В–∞–і–Є–Є —А–∞–Ј–ї–Њ–ґ–µ–љ–Є—П (–њ–Є–ї–Є—В—Б—П –Ї–∞–Ї —Б–ї–µ–≥–Ї–∞ —В—А—Г—Е–ї—П–≤–Њ–µ –і–µ—А–µ–≤–Њ), –љ–Њ —Б–Њ—Е—А–∞–љ—П–µ—В —Б—В—А—Г–Ї—В—Г—А—Г –≥–Њ–і–Є—З–љ—Л—Е –Ї–Њ–ї–µ—Ж, –Њ—Б–Њ–±–µ–љ–љ–Њ –≤ –Є–Ј–≤–ї–µ—З—С–љ–љ–Њ–є –њ–Њ–≥—А–µ–±—С–љ–љ–Њ–є –Є –Њ—В—В–∞—П–≤—И–µ–є —З–∞—Б—В–Є.
–Т –і–∞–љ–љ–Њ–Љ –Љ–µ—Б—В–µ –љ–∞ —Б–Ї–ї–Њ–љ–µ –љ–∞—Б—З–Є—В—Л–≤–∞–µ—В—Б—П –µ—Й—С –і–≤–µ —В–∞–Ї–Є—Е –њ–∞—З–Ї–Є –њ–µ—А–µ—Б–ї–∞–Є–≤–∞–љ–Є—П, —Б—К–µ–Ј–ґ–∞—О—Й–Є—Е –≤–љ–Є–Ј –њ–Њ –Њ—Б—Л–њ–љ–Њ–Љ—Г —Б–Ї–ї–Њ–љ—Г. –Ф–Њ –±—А–Њ–≤–Ї–Є –љ–∞ –≤—Л—Б–Њ—В–µ 51,7 –Љ –љ–∞–і —Г—А–Њ–≤–љ–µ–Љ –Љ–Њ—А—П –љ–∞ –Њ—Б—Л–њ–љ–Њ–Љ —Б–Ї–ї–Њ–љ–µ –≤—Б—В—А–µ—З–∞—О—В—Б—П –Ї–Њ—Б—В–Є –Љ–Њ—А—Б–Ї–Є—Е –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е, –Ї–Њ—В–Њ—А—Л–µ –њ–Њ–њ–∞–і–∞—О—В –љ–∞ –і–љ–Њ –і–Њ–ї–Є–љ—Л –Є–Љ–µ–љ–љ–Њ –Є–Ј —Н—В–Є—Е –≥—А–∞–≤–Є–є–љ–Є–Ї–Њ–≤ –Є –≥–∞–ї–µ—З–љ–Є–Ї–Њ–≤.
–Т–Њ–Ј–≤—А–∞—Й–µ–љ–Є–µ –Ї –Њ–±–љ–∞–ґ–µ–љ–Є—О I, –Є–Ј –Ї–Њ—В–Њ—А–Њ–≥–Њ 9 –∞–≤–≥—Г—Б—В–∞ –±—Л–ї–Є –Њ—В–Њ–±—А–∞–љ—Л –∞–ї–µ–≤—А–Є—В—Л –і–ї—П –≠–Я–† –∞–љ–∞–ї–Є–Ј–∞. –Я–Њ–і–Њ—И–≤–∞ —Н—В–Њ–≥–Њ –Њ–±–љ–∞–ґ–µ–љ–Є—П, –≤—Б–Ї—А—Л—В–Њ–≥–Њ —А—Г—З—М—С–Љ –њ–Њ –њ—А–∞–≤–Њ–Љ—Г —Б–Ї–ї–Њ–љ—Г –і–Њ–ї–Є–љ—Л, –ї–µ–ґ–Є—В –љ–∞ –≤—Л—Б–Њ—В–µ 9,5 –Љ –љ–∞–і —Г—А–Њ–≤–љ–µ–Љ –Љ–Њ—А—П. –Ґ–Њ–ї—Й–∞ –∞–ї–µ–≤—А–Є—В–Њ–≤ –Є–Љ–µ–µ—В –≤–Є–і–Є–Љ—Г—О –Љ–Њ—Й–љ–Њ—Б—В—М 10 –Љ. –Т –Њ–±—А–∞—Й—С–љ–љ–Њ–є –Ї —А—Г—З—М—О —Б—В–µ–љ–Ї–µ —Г–≥–Њ–ї —Б–Ї–ї–Њ–љ–∞ –њ—А–∞–Ї—В–Є—З–µ—Б–Ї–Є –≤–µ—А—В–Є–Ї–∞–ї—М–љ—Л–є. –Ъ —А–∞–Ј—А–µ–Ј—Г –Љ–Њ–ґ–љ–Њ –±—Л–ї–Њ –њ–Њ–і–Њ–±—А–∞—В—М—Б—П –њ–Њ –і–Њ–ї–Є–љ–Ї–µ –Љ–∞–ї–µ–љ—М–Ї–Њ–≥–Њ –Њ–≤—А–∞–ґ–Ї–∞, –Њ—В—Е–Њ–і—П—Й–µ–≥–Њ –Њ—В —А—Г—З—М—П –Ї –Ј–∞–њ–∞–і—Г.
–Т—Б—П —В–Њ–ї—Й–∞ –∞–ї–µ–≤—А–Є—В–Њ–≤ –±–Њ–ї–µ–µ –Є–ї–Є –Љ–µ–љ–µ–µ –Њ–і–љ–Њ—А–Њ–і–љ–∞—П –њ—А–µ–і—Б—В–∞–≤–ї–µ–љ–∞ –њ–µ—А–µ—Б–ї–∞–Є–≤–∞–љ–Є–µ–Љ —Б–µ—А—Л—Е –Є –ґ–µ–ї—В–Њ–≤–∞—В–Њ-—Б–µ—А—Л—Е —Б–ї–Њ—С–≤ –∞–ї–µ–≤—А–Є—В–Њ–≤. –Ш–љ–Њ–≥–і–∞ –≤ —В–Њ–ї—Й–µ —Б–Њ–і–µ—А–ґ–∞—В—Б—П –њ—А–Њ—Б–ї–Њ–є–Ї–Є-–ї–Є–љ–Ј—Л –≥–∞–ї–µ—З–љ–Є–Ї–Њ–≤ —В–Њ–ї—Й–Є–љ–Њ–є –і–Њ 1вАУ2 —Б–Љ (–Љ–µ–ї–Ї–∞—П –≥–∞–ї—М–Ї–∞ –Є –≥—А–∞–≤–Є–є). –Ю–±—А–∞–Ј–µ—Ж 3298/14 –∞–ї–µ–≤—А–Є—В–Њ–≤ –Њ—В–Њ–±—А–∞–љ —Б –≥–ї—Г–±–Є–љ—Л 1 –Љ –љ–Є–ґ–µ –±—А–Њ–≤–Ї–Є –Њ–±–љ–∞–ґ–µ–љ–Є—П (18, 5 –Љ –љ–∞–і —Г.–Љ.) –Т 1 –Љ –љ–Є–ґ–µ –±—А–Њ–≤–Ї–Є –≤ –∞–ї–µ–≤—А–Є—В–∞—Е –њ–Њ–њ–∞–і–∞—О—В—Б—П —Ж–µ–ї—Л–µ —А–∞–Ї–Њ–≤–Є–љ—Л –Љ–Њ–ї–ї—О—Б–Ї–Њ–≤ –Є –Є—Е –Њ—Б–Ї–Њ–ї–Ї–Є, –Њ—З–µ–љ—М —В–Њ–љ–Ї–Є–µ, —А–∞–Ј–≤–∞–ї–Є–≤–∞—О—Й–Є–µ—Б—П –њ—А–Є –Є–Ј–≤–ї–µ—З–µ–љ–Є–Є –Є–Ј –Њ—Б–∞–і–Ї–∞. –Х—Б—В—М —А–∞–Ї–Њ–≤–Є–љ—Л –і–≤—Г—Б—В–≤–Њ—А—З–∞—В—Л—Е –Љ–Њ–ї–ї—О—Б–Ї–Њ–≤ –Є –≥–∞—Б—В—А–Њ–њ–Њ–і. –Т –≥–Њ—А–Є–Ј–Њ–љ—В–µ 18, 5 –Љ –љ–∞–і —Г—А–Њ–≤–љ–µ–Љ –Љ–Њ—А—П —Б–Њ–±—А–∞–љ—Л —В–Њ–љ–Ї–Є–µ —А–∞–Ї–Њ–≤–Є–љ—Л –і–≤—Г—Б—В–≤–Њ—А—З–∞—В—Л—Е –Љ–Њ–ї–ї—О—Б–Ї–Њ–≤ –≤ –њ—А–Њ–±–Є—А–Ї—Г вАУ –Њ–±—А–∞–Ј–µ—Ж 3298/15.
–Э–∞ –љ–µ–≤—Б–Ї—А—Л—В–Њ–є –њ–Њ–≤–µ—А—Е–љ–Њ—Б—В–Є –Њ–±–љ–∞–ґ–µ–љ–Є—П –µ—Б—В—М –±–µ–ї—Л–µ –Є —Б–µ—А—Л–µ –≤—Л—Ж–≤–µ—В—Л —Б–Њ–ї–µ–є –њ–Њ –≤–Ї—Г—Б—Г –њ–Њ–≤–∞—А–µ–љ–љ–Њ–є —Б–Њ–ї–Є вАУ –Њ–±—А. 3298/16 —Б –≥–Њ—А–Є–Ј–Њ–љ—В–∞ 18,5 –Љ. –Э–∞ –≤—Л—Б–Њ—В–µ 17,5 –Љ –љ–∞–є–і–µ–љ–∞ —А–∞–Ї–Њ–≤–Є–љ–∞ –≥–∞—Б—В—А–Њ–њ–Њ–і—Л –і–Є–∞–Љ–µ—В—А–Њ–Љ 1 —Б–Љ, —Е—А—Г–њ–Ї–∞—П вАУ –Њ—В–Њ–±—А–∞–љ–∞ –≤ –њ–ї–∞—Б—В–Є–Ї–Њ–≤—Г—О –њ—А–Њ–±–Є—А–Ї—Г вАУ –Њ–±—А. 3298/17. –≠—В–Њ –≥–Њ—А–Є–Ј–Њ–љ—В, –Є–Ј –Ї–Њ—В–Њ—А–Њ–≥–Њ 9 –∞–≤–≥—Г—Б—В–∞ –±—Л–ї –Њ—В–Њ–±—А–∞–љ –Њ–±—А–∞–Ј–µ—Ж 3298/1. –Ю—В—Б—О–і–∞ –ґ–µ –Њ—В–Њ–±—А–∞–љ –Њ–±—А–∞–Ј–µ—Ж –∞–ї–µ–≤—А–Є—В–Њ–≤ 3298/18 –≤ —В—А—Г–±–Ї—Г –і–ї—П –њ—А–Њ–≤–µ–і–µ–љ–Є—П –Ю–°–Ы-–∞–љ–∞–ї–Є–Ј–∞ –≤–Њ–Ј—А–∞—Б—В–∞. –° –≤—Л—Б–Њ—В—Л 16 –Љ –Њ—В–Њ–±—А–∞–љ—Л —Б–Њ–ї–Є, –Ї–Њ—В–Њ—А—Л–µ –њ—А–Њ—Б–ї–µ–ґ–Є–≤–∞—О—В—Б—П –≤ —А–∞–Ј—А–µ–Ј–µ –Ї–∞–Ї –њ—А–Њ—Б–ї–Њ–є —В–Њ–ї—Й–Є–љ–Њ–є –і–Њ 1 —Б–Љ. –°–Њ–ї–Є –њ—А–µ–і—Б—В–∞–≤–ї–µ–љ—Л –њ—А–Њ–Ј—А–∞—З–љ—Л–Љ–Є –Ї—А–Є—Б—В–∞–ї–ї–∞–Љ–Є —Б —Б–Њ–ї—С–љ—Л–Љ –Є –њ—А–Њ—В–Є–≤–љ—Л–Љ –≤–Ї—Г—Б–Њ–Љ. –Ш–Ј –љ–Є—Е –≤–Ј—П—В –Њ–±—А–∞–Ј–µ—Ж 3298/19. –Э–Є–ґ–љ—П—П —З–∞—Б—В—М –Њ–±–љ–∞–ґ–µ–љ–Є—П —А–∞—Б—Б–Љ–Њ—В—А–µ–љ–∞ —Б–Њ —Б—В–Њ—А–Њ–љ—Л –≤–µ—А—В–Є–Ї–∞–ї—М–љ–Њ–є —Н—А–Њ–Ј–Є–Њ–љ–љ–Њ–є —Б—В–µ–љ–Ї–Є —А—Г—З—М—П. –Ю–љ–∞ —Е–∞—А–∞–Ї—В–µ—А–љ–∞ –њ–µ—А–µ—Б–ї–∞–Є–≤–∞–љ–Є–µ–Љ —Б–µ—А—Л—Е –∞–ї–µ–≤—А–Є—В–Њ–≤ –±–Њ–ї—М—И–µ–є —З–∞—Б—В—М—О (—Б–ї–Њ–є–Ї–Є –њ–Њ 3вАУ10 —Б–Љ) –Є –њ—А–Њ—Б–ї–Њ—П–Љ–Є –∞–ї–µ–≤—А–Є—В–Њ–Љ —А—Л–ґ–µ–≥–Њ —Ж–≤–µ—В–∞ –њ–Њ 1вАУ2 —Б–Љ —В–Њ–ї—Й–Є–љ–Њ–є. –Э–∞ –≤—Л—Б–Њ—В–µ 2 –Љ –љ–∞–і –њ–Њ–і–Њ—И–≤–Њ–є –Њ–±–љ–∞–ґ–µ–љ–Є—П (11,5 –Љ –љ–∞–і —Г.–Љ.) –Ј–∞–ї–µ–≥–∞–µ—В –њ—А–Њ—Б–ї–Њ–є –Љ–µ–ї–Ї–Њ–≥–Њ –≥–∞–ї–µ—З–љ–Є–Ї–∞ —В–Њ–ї—Й–Є–љ–Њ–є 15вАУ20 —Б–Љ. –Т –њ–Њ–і–Њ—И–≤–µ –≥–∞–ї–µ—З–љ–Є–Ї–∞ –µ—Б—В—М –њ—А–Њ—Б–ї–Њ–є–Ї–Є –≥—А–∞–≤–Є—П –Є –њ–µ—Б–Ї–∞, –Є–Ј –Ї–Њ—В–Њ—А—Л—Е –≤–Ј—П—В –Њ–±—А–∞–Ј–µ—Ж 3298/20. –У–∞–ї—М–Ї–Є —А–∞—Б–њ–Њ–ї–Њ–ґ–µ–љ—Л –њ—А–Њ—Б–ї–Њ—П–Љ–Є, –њ–∞–і–∞—О—Й–Є–Љ–Є –≤ —А–∞–Ј–љ—Л–µ —Б—В–Њ—А–Њ–љ—Л –њ–Њ–і –±–Њ–ї—М—И–Є–Љ–Є —Г–≥–ї–∞–Љ–Є. –Ш–Ј –Њ—Б–љ–Њ–≤–∞–љ–Є—П —Г—Б—В—Г–њ–∞ —А–∞–Ј–Љ—Л–≤–∞ —Б –≤—Л—Б–Њ—В—Л 0,4 –Љ (9,9 –Љ –љ–∞–і —Г.–Љ.) –≤–Ј—П—В –Њ–±—А–∞–Ј–µ—Ж –∞–ї–µ–≤—А–Є—В–∞ 3298/21. –Т —Б—В–µ–љ–Ї–µ –љ–∞ –≤—Л—Б–Њ—В–µ –њ—А–Є–Љ–µ—А–љ–Њ 5 –Є 6,5 –Љ –љ–∞–і –Њ—Б–љ–Њ–≤–∞–љ–Є–µ–Љ (14,5 –Є 16 –Љ –љ–∞–і —Г.–Љ.) –≤–Є–і–љ—Л —В–Њ–љ–Ї–Є–µ –њ—А–Њ—Б–ї–Њ–є–Ї–Є —Б–Њ–ї–µ–є, –Ї–Њ—В–Њ—А—Л–µ –Њ—В–Њ–±—А–∞–љ—Л –Є–Ј —Б—В–µ–љ–Ї–Є –≤ —А–∞—Б–њ–∞–і–Ї–µ (–Њ–±—А. 3298/19).
–Т —Ж–µ–ї–Њ–Љ —А–∞–Ј—А–µ–Ј—Л –њ–Њ–Ї–∞–Ј–∞–ї–Є, —З—В–Њ –Љ–Њ—А—Б–Ї–Є–µ –Њ—В–ї–Њ–ґ–µ–љ–Є—П —Б–Њ—Б—В–Њ—П—В –Є–Ј 3 –њ–∞—З–µ–Ї –Њ—Б–∞–і–Ї–Њ–≤. –Я–Њ –≤—Л—Б–Њ—В–µ —Б–∞–Љ–∞—П –љ–Є–Ј–Ї–∞—П –њ–∞—З–Ї–∞ –Њ—В–ї–Њ–ґ–µ–љ–Є–є вАУ –∞–ї–µ–≤—А–Є—В–Њ–≤–∞—П, —А–∞—Б–њ–Њ–ї–∞–≥–∞–µ—В—Б—П –±–ї–Є–ґ–µ –Ї –Љ–Њ—А—О (9,5вАУ20 –Љ). –°—А–µ–і–љ—П—П вАУ —З—С—А–љ—Л–µ –≥–ї–Є–љ—Л –љ–∞ –≤—Л—Б–Њ—В–µ 25вАУ33,7 –Љ, –≤—Л—Б–Њ–Ї–∞—П вАУ –≥—А–∞–≤–Є–є–љ–Њ-–≥–∞–ї–µ—З–љ–∞—П —В–Њ–ї—Й–∞ (33.7вАУ50 –Љ). –Т—Л—Б–Њ—В—Л –Њ–њ—А–µ–і–µ–ї–µ–љ—Л —Н–Ї–ї–Є–Љ–µ—В—А–Є—А–Њ–≤–∞–љ–Є–µ–Љ –Є –њ–Њ –і–∞–љ–љ—Л–Љ –њ—А–Є–±–Њ—А–∞ GPS. –Ч–љ–∞—З–Є—В –Є—Е —В–Њ—З–љ–Њ—Б—В—М –ї–µ–ґ–Є—В –≤ –њ—А–µ–і–µ–ї–∞—Е 1вАУ2 –Љ.
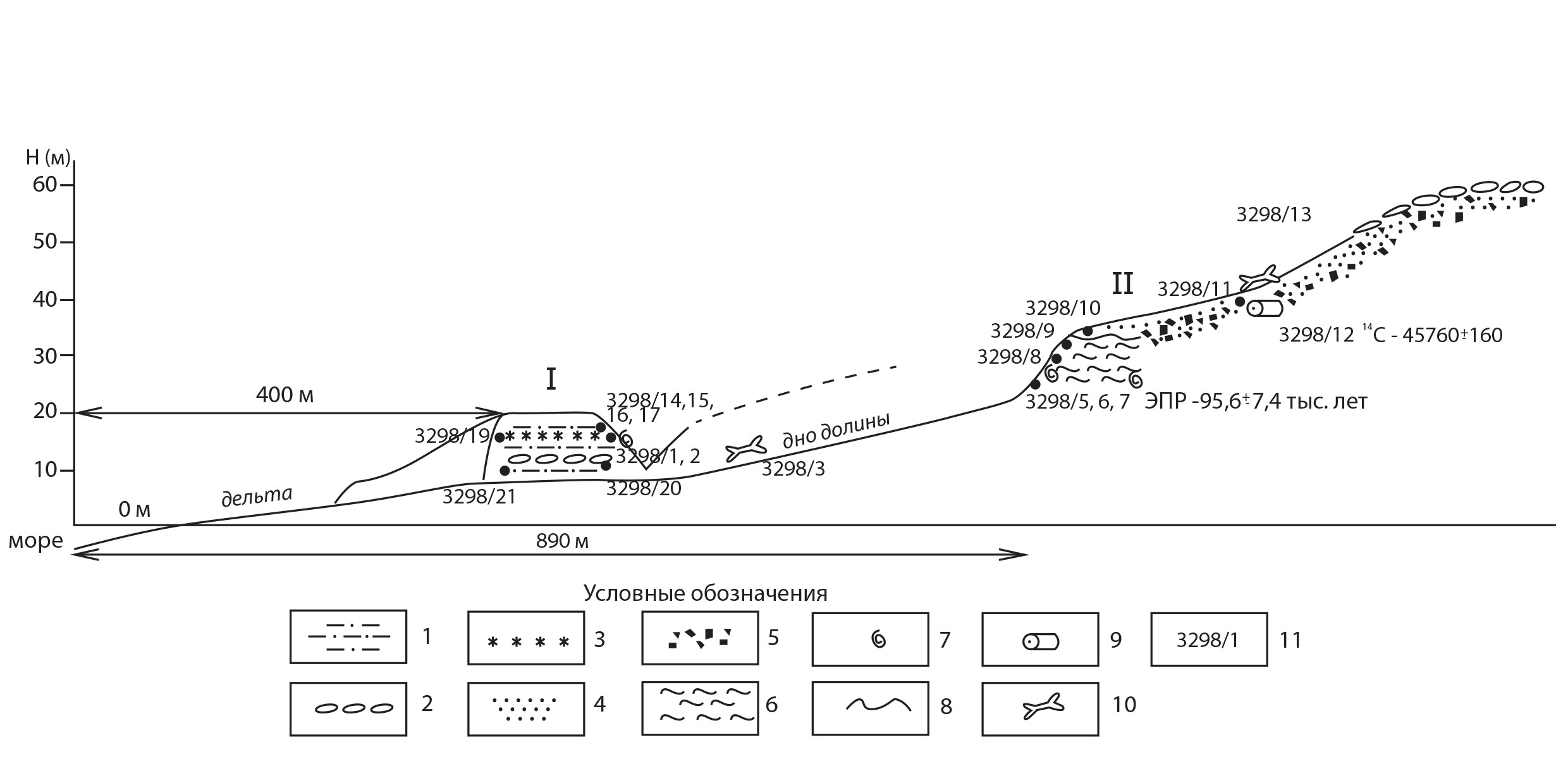
–†–Є—Б. 1 –°—Е–µ–Љ–∞ –Њ–±–љ–∞–ґ–µ–љ–Є—П 3298, –Є–Ј—Г—З–µ–љ–љ–Њ–≥–Њ –≤ –і–Њ–ї–Є–љ–µ —А–µ–Ї–Є, –≤–њ–∞–і–∞—О—Й–µ–є –≤ –Ј–∞–ї–Є–≤ –Р—Е–Љ–∞—В–Њ–≤–∞ —Б –Ј–∞–њ–∞–і–љ–Њ–≥–Њ —Б–Ї–ї–Њ–љ–∞. –£—Б–ї–Њ–≤–љ—Л–µ –Њ–±–Њ–Ј–љ–∞—З–µ–љ–Є—П: 1 вАУ –∞–ї–µ–≤—А–Є—В, 2 вАУ –≥–∞–ї—М–Ї–∞, 3 вАУ –њ—А–Њ—Б–ї–Њ–є —Б–Њ–ї–Є, 4 вАУ –њ–µ—Б–Њ–Ї, 5 вАУ –≥—А–∞–≤–Є–є, 6 вАУ –≥–ї–Є–љ–∞, 7 вАУ—А–∞–Ї–Њ–≤–Є–љ–∞ –Љ–Њ–ї–ї—О—Б–Ї–∞, 8 вАУ –≥—А–∞–љ–Є—Ж–∞ —Б—В—А–∞—В–Є–≥—А–∞—Д–Є—З–µ—Б–Ї–Њ–≥–Њ –љ–µ—Б–Њ–≥–ї–∞—Б–Є—П, 9- –њ–ї–∞–≤–љ–Є–Ї –і—А–µ–≤–µ—Б–Є–љ—Л, 10 вАУ –Ї–Њ—Б—В—М –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–µ–≥–Њ, 11 вАУ –љ–Њ–Љ–µ—А –Њ–±—А–∞–Ј—Ж–∞

–†–Є—Б. 2 –†–∞–Ј—А–µ–Ј (I) –Љ–Њ—А—Б–Ї–Є—Е –∞–ї–µ–≤—А–Є—В–Њ–≤ –≤ 400 –Љ –Њ—В –Ї—А–∞—П –і–µ–ї—М—В—Л –±–µ–Ј—Л–Љ—П–љ–љ–Њ–є —А–µ–Ї–Є, –≤–њ–∞–і–∞—О—Й–µ–є –≤ –Ј–∞–ї–Є–≤ –Р—Е–Љ–∞—В–Њ–≤–∞

–†–Є—Б. 3 –•–Њ—А–Њ—И–Њ –Њ–Ї–∞—В–∞–љ–љ–∞—П –≥–∞–ї—М–Ї–∞ –љ–∞ –њ–Њ–≤–µ—А—Е–љ–Њ—Б—В–Є –Љ–Њ—А—Б–Ї–Њ–є —В–µ—А—А–∞—Б—Л –≤—Л—Б–Њ—В–Њ–є 50вАУ60 –Љ –љ–∞ –Ј–∞–њ–∞–і–љ–Њ–Љ —Б–Ї–ї–Њ–љ–µ –Ј–∞–ї–Є–≤–∞ –Р—Е–Љ–∞—В–Њ–≤–∞

–†–Є—Б. 4 –†–∞–Ї–Њ–≤–Є–љ—Л –і–≤—Г—Б—В–≤–Њ—А—З–∞—В—Л—Е –Љ–Њ–ї–ї—О—Б–Ї–Њ–≤ –≤ —З—С—А–љ—Л—Е –≥–ї–Є–љ–∞—Е –≤ –Њ—Б–љ–Њ–≤–∞–љ–Є–Є —А–∞–Ј—А–µ–Ј–∞ II

–†–Є—Б. 5 –†–∞–Ј–Љ—Л–≤ —А—Г—З—М—С–Љ —З—С—А–љ—Л—Е –≥–ї–Є–љ —Б –±–∞–љ–Ї–Њ–є –і–≤—Г—Б—В–≤–Њ—А—З–∞—В—Л—Е –Љ–Њ–ї–ї—О—Б–Ї–Њ–≤ (—А–∞–Ј—А–µ–Ј II)
–Ф–Є–∞—В–Њ–Љ–Њ–≤—Л–є –∞–љ–∞–ї–Є–Ј 12 –Њ–±—А–∞–Ј—Ж–Њ–≤ –Њ–±–љ–∞–ґ–µ–љ–Є—П 3298, –≤—Л–њ–Њ–ї–љ–µ–љ –Љ–Є–Ї—А–Њ–њ–∞–ї–µ–љ—В–Њ–ї–Њ–≥–Њ–Љ –Ч.–Т. –Я—Г—И–Є–љ–Њ–є (–Р–Р–Э–Ш–Ш, –Т–Э–Ш–Ш–Ю–Ї–µ–∞–љ–≥–µ–Њ–ї–Њ–≥–Є—П). –Ъ–Њ–ї–Є—З–µ—Б—В–≤–Њ —Б—В–≤–Њ—А–Њ–Ї –і–Є–∞—В–Њ–Љ–µ–є –љ–µ–Ј–љ–∞—З–Є—В–µ–ї—М–љ–Њ, —Б–Њ—Е—А–∞–љ–љ–Њ—Б—В—М —Б—В–≤–Њ—А–Њ–Ї –Њ—З–µ–љ—М –≤–∞—А—М–Є—А—Г–µ—В: –≤—Б—В—А–µ—З–µ–љ—Л —Ж–µ–ї—Л–µ —Б—В–≤–Њ—А–Ї–Є, –љ–Њ –Љ–љ–Њ–≥–Њ –Њ–±–ї–Њ–Љ–Њ—З–љ–Њ–≥–Њ –Љ–∞—В–µ—А–Є–∞–ї–∞, –≤ –љ–µ–Ї–Њ—В–Њ—А—Л—Е –Њ–±—А–∞–Ј—Ж–∞—Е —Г—Б—В–∞–љ–Њ–≤–ї–µ–љ—Л —В–Њ–ї—М–Ї–Њ —Д—А–∞–≥–Љ–µ–љ—В—Л —Б—В–≤–Њ—А–Њ–Ї –і–Є–∞—В–Њ–Љ–µ–є. –Т –і–Є–∞—В–Њ–Љ–Њ–≤—Л—Е –Ї–Њ–Љ–њ–ї–µ–Ї—Б–∞—Е —Г—Б—В–∞–љ–Њ–≤–ї–µ–љ–Њ 40 –≤–Є–і–Њ–≤ –і–Є–∞—В–Њ–Љ–µ–є. –Т –Є—Е —Б–Њ—Б—В–∞–≤ –≤—Е–Њ–і—П—В –љ–µ—А–Є—В–Є—З–µ—Б–Ї–Є–µ –Є –Њ–Ї–µ–∞–љ–Є—З–µ—Б–Ї–Є–µ –і–Є–∞—В–Њ–Љ–µ–Є вАУ 13 –≤–Є–і–Њ–≤, –±–µ–љ—В–Є—З–µ—Б–Ї–Є—Е —Б—Г–±–ї–Є—В–Њ—А–∞–ї—М–љ—Л—Е - 26 –≤–Є–і–Њ–≤. (–Ґ–∞–±–ї–Є—Ж–∞ 1). ¬Ђ–Х–і–Є–љ–Є—З–љ–Њ¬ї –≤—Б—В—А–µ—З–µ–љ –і–Є–∞—В–Њ–Љ–Њ–≤—Л–є –≤–Є–і Rhabdonema cf. japonicum, –Є–Ј–≤–µ—Б—В–љ—Л–є —В–Њ–ї—М–Ї–Њ –≤ –Њ—Б–∞–і–Ї–∞—Е –љ–µ–Њ–≥–µ–љ–Њ–≤–Њ–≥–Њ –≤–Њ–Ј—А–∞—Б—В–∞. –Ґ–∞–Ї–ґ–µ –≤ –Є—Б—Б–ї–µ–і—Г–µ–Љ—Л—Е –Њ—В–ї–Њ–ґ–µ–љ–Є—П—Е –Њ–±–љ–∞—А—Г–ґ–µ–љ—Л —Ж–Є—Б—В—Л –Ј–Њ–ї–Њ—В–Є—Б—В—Л—Е –≤–Њ–і–Њ—А–Њ—Б–ї–µ–є (Chrysophycean cysts).
–У–Њ—Б–њ–Њ–і—Б—В–≤—Г—О—Й—Г—О –≥—А—Г–њ–њ—Г –і–Є–∞—В–Њ–Љ–µ–є –≤ –Є–Ј—Г—З–µ–љ–љ—Л—Е –Њ—В–ї–Њ–ґ–µ–љ–Є—П—Е —Б–Њ—Б—В–∞–≤–ї—П—О—В –Љ–Њ—А—Б–Ї–Є–µ –і–Є–∞—В–Њ–Љ–Њ–≤—Л–µ –≤–Њ–і–Њ—А–Њ—Б–ї–Є, —З–Є—Б–ї–Њ –Ї–Њ—В–Њ—А—Л—Е —Б–Њ—Б—В–∞–≤–ї—П–µ—В 28 –≤–Є–і–Њ–≤ (Actinocyclus sp., Chaetoceros sp., Diploneis subcincta, Thalassiosira gravida, Trachyneis aspera –Є –і—А—Г–≥–Є–µ). –Т –і–Є–∞—В–Њ–Љ–Њ–≤—Л—Е –Ї–Њ–Љ–њ–ї–µ–Ї—Б–∞—Е —З–∞—Б—В–Њ –≤—Б—В—А–µ—З–µ–љ—Л —Б–Њ–ї–Њ–љ–Њ–≤–∞—В–Њ–≤–Њ–і–љ–Њ-–Љ–Њ—А—Б–Ї–Є–µ –≤–Є–і—Л Cocconeis scutellum, Diploneis smithii, Grammatophora angulosa –Є –і—А—Г–≥–Є–µ, —Б–Њ–ї–Њ–љ–Њ–≤–∞—В–Њ–≤–Њ–і–љ—Л–µ –њ—А–µ–і—Б—В–∞–≤–ї–µ–љ—Л ¬Ђ–µ–і–Є–љ–Є—З–љ–Њ¬ї Bacillaria socialis, Thalassiosira baltica, —В–∞–Ї–ґ–µ –Њ–±–љ–∞—А—Г–ґ–µ–љ—Л –њ—А–µ—Б–љ–Њ–≤–Њ–і–љ—Л–µ Fragilaria spp., Pinnularia obscura, Ulnaria ulna –Є –і—А—Г–≥–Є–µ.
–С–Њ–ї—М—И–Є–љ—Б—В–≤–Њ —Г—Б—В–∞–љ–Њ–≤–ї–µ–љ–љ—Л—Е –і–Є–∞—В–Њ–Љ–Њ–≤—Л—Е –≤–Њ–і–Њ—А–Њ—Б–ї–µ–є —П–≤–ї—П—О—В—Б—П –Ї–Њ—Б–Љ–Њ–њ–Њ–ї–Є—В–∞–Љ–Є –Є –њ—А–∞–Ї—В–Є—З–µ—Б–Ї–Є –≤—Б–µ –Є–Ј–≤–µ—Б—В–љ—Л –≤ —Б–Њ–≤—А–µ–Љ–µ–љ–љ—Л—Е –і–Є–∞—В–Њ–Љ–Њ–≤—Л—Е –±–Є–Њ—Ж–µ–љ–Њ–Ј–∞—Е –Є –≤ –і–Є–∞—В–Њ–Љ–Њ–≤—Л—Е –Ї–Њ–Љ–њ–ї–µ–Ї—Б–∞—Е –Є–Ј –≥–Њ–ї–Њ—Ж–µ–љ–Њ–≤—Л—Е –Њ—В–ї–Њ–ґ–µ–љ–Є–є –Љ–Њ—А–µ–є –Т–Њ—Б—В–Њ—З–љ–Њ–є –Р—А–Ї—В–Є–Ї–Є (–Я–Њ–ї—П–Ї–Њ–≤–∞ –Х. –Ш., 1997; Cremer H., 1999; –¶–Њ–є –Ш. –С., –Ю–±—А–µ–Ј–Ї–Њ–≤–∞ –Ь. –°., 2017).
–Т –љ–Є–ґ–љ–µ–є –∞–ї–µ–≤—А–Є—В–Њ–≤–Њ–є –њ–∞—З–Ї–µ –Њ—В–Њ–±—А–∞–љ—Л –Њ–±—А–∞–Ј—Ж—Л 3298/21, 3298/20, 3298/2, 3298/4, 3298/14, 3298/16, 3298/19. –Т —Б–∞–Љ–Њ–Љ –љ–Є–ґ–љ–µ–Љ —Б–ї–Њ–µ –∞–ї–µ–≤—А–Є—В–Њ–≤ –Њ–±—А–∞–Ј–µ—Ж 3298/21 —Г—Б—В–∞–љ–Њ–≤–ї–µ–љ—Л –і–Є–∞—В–Њ–Љ–Њ–≤—Л–µ –Ї–Њ–Љ–њ–ї–µ–Ї—Б—Л in situ, —Б–Њ—Б—В–Њ—П—Й–Є–µ –Є–Ј –љ–µ–Љ–љ–Њ–≥–Њ—З–Є—Б–ї–µ–љ–љ—Л—Е —Б—В–≤–Њ—А–Њ–Ї –Љ–Њ—А—Б–Ї–Є—Е –і–Є–∞—В–Њ–Љ–Њ–≤—Л—Е –≤–Њ–і–Њ—А–Њ—Б–ї–µ–є, —В–∞–Ї–ґ–µ –њ—А–Є—Б—Г—В—Б—В–≤—Г—О—В —Ж–Є—Б—В—Л –Ј–Њ–ї–Њ—В–Є—Б—В—Л—Е –≤–Њ–і–Њ—А–Њ—Б–ї–µ–є. –Ф–Є–∞—В–Њ–Љ–µ–Є –њ—А–µ–і—Б—В–∞–≤–ї–µ–љ—Л –Ї–∞–Ї –Њ—В–љ–Њ—Б–Є—В–µ–ї—М–љ–Њ –≥—А—Г–±–Њ–Њ–Ї—А–µ–Љ–љ–µ–љ–љ—Л–Љ–Є —Б—В–≤–Њ—А–Ї–∞–Љ–Є –Љ–Њ—А—Б–Ї–Є—Е –і–Є–∞—В–Њ–Љ–Њ–≤—Л—Е, —В–∞–Ї –Є —В–Њ–љ–Ї–Њ–Њ–Ї—А–µ–Љ–µ–љ–µ–љ–љ—Л–Љ–Є —Б—В–≤–Њ—А–Ї–∞–Љ–Є, –Љ–∞–ї–µ–љ—М–Ї–Њ–≥–Њ —А–∞–Ј–Љ–µ—А–∞ (Fragilaria spp.; –і–ї–Є–љ–∞ –Њ–Ї–Њ–ї–Њ 15 –Љ–Є–Ї—А–Њ–љ) —Е–Њ—А–Њ—И–µ–є —Б–Њ—Е—А–∞–љ–љ–Њ—Б—В–Є. –Т–µ—А–Њ—П—В–љ–Њ, —Н—В–Є —В–Њ–љ–Ї–Њ–Њ–Ї—А–µ–Љ–љ–µ–љ–љ—Л–µ —Б—В–≤–Њ—А–Ї–Є –њ—А–µ—Б–љ–Њ–≤–Њ–і–љ—Л—Е –і–Є–∞—В–Њ–Љ–µ–є –±—Л–ї–Є –Ј–∞–љ–µ—Б–µ–љ—Л –≤ –Њ—В–ї–Њ–ґ–µ–љ–Є—П –њ–Њ–Ј–і–љ–µ–µ, –њ–Њ—Б–Ї–Њ–ї—М–Ї—Г —Б—В–≤–Њ—А–Ї–Є –Љ–Њ—А—Б–Ї–Є—Е –і–Є–∞—В–Њ–Љ–µ–є –≤—Б—В—А–µ—З–µ–љ—Л –Є –≤ –Њ–±–ї–Њ–Љ–Њ—З–љ–Њ–Љ —Б–Њ—Б—В–Њ—П–љ–Є–Є –Є –Љ–∞–ї–Њ, –≤–µ—А–Њ—П—В–љ–Њ, —З—В–Њ–±—Л —В–∞–Ї–Є–µ ¬Ђ—В–Њ–љ–Ї–Є–µ¬ї —Б—В–≤–Њ—А–Ї–Є –њ—А–µ—Б–љ–Њ–≤–Њ–і–љ—Л—Е –Љ–Њ–≥–ї–Є —Б–Њ—Е—А–∞–љ–Є—В—М—Б—П —Ж–µ–ї—Л–Љ–Є.
–Т –Њ–±—А–∞–Ј—Ж–µ 3298/21 –љ–∞–Є–±–Њ–ї–µ–µ —З–∞—Б—В–Њ –≤—Б—В—А–µ—З–µ–љ—Л –і–Є–∞—В–Њ–Љ–Њ–≤—Л–µ –≤–Њ–і–Њ—А–Њ—Б–ї–Є:
–љ–µ—А–Є—В–Є—З–µ—Б–Ї–Є–µ –њ–ї–∞–љ–Ї—В–Њ–љ–љ—Л–µ A—Бtinocyclus sp., Bacterosira fragilis, Chaetoceros sp., Thalassiosira gravida, –Њ–Ї–µ–∞–љ–Є—З–µ—Б–Ї–∞—П Rhizosolenia styliformis; –ї–µ–і–Њ–≤–Њ-–Љ–Њ—А—Б–Ї–Є–µ –њ–ї–∞–љ–Ї—В–Њ–љ–љ—Л–µ Fragilariopsis oceanica; –±–µ–љ—В–Њ—Б–љ—Л–µ Amphora sp., Cocconeis costata, C. scutellum, Grammatophora angulosa, Diploneis smithii, Rhabdonema sp., Trachyneis aspera –Є –і—А—Г–≥–Є–µ. –Я—А–Є—Б—Г—В—Б—В–≤–Є–µ —Е–Њ—В—П –Є –љ–µ–Љ–љ–Њ–≥–Њ—З–Є—Б–ї–µ–љ–љ—Л—Е —Б—В–≤–Њ—А–Њ–Ї –њ–ї–∞–љ–Ї—В–Њ–љ–љ—Л—Е –і–Є–∞—В–Њ–Љ–Њ–≤—Л—Е –њ–Њ–Ј–≤–Њ–ї—П–µ—В —Б–і–µ–ї–∞—В—М –≤—Л–≤–Њ–і –Њ —Д–Њ—А–Љ–Є—А–Њ–≤–∞–љ–Є–Є –Њ—В–ї–Њ–ґ–µ–љ–Є–є, –≤–Љ–µ—Й–∞—О—Й–Є—Е —Н—В–Њ—В –і–Є–∞—В–Њ–Љ–Њ–≤—Л–є –Ї–Њ–Љ–њ–ї–µ–Ї—Б, –≤ –њ—А–Є–±—А–µ–ґ–љ—Л—Е –Њ—В–љ–Њ—Б–Є—В–µ–ї—М–љ–Њ –≥–ї—Г–±–Њ–Ї–Є—Е —Г—Б–ї–Њ–≤–Є—П—Е –Љ–Њ—А—Б–Ї–Њ–≥–Њ –±–∞—Б—Б–µ–є–љ–∞.
–Т –Њ–±—А–∞–Ј—Ж–µ 3298/20 –Є–Ј –њ–µ—Б—З–∞–љ–Њ-–≥–∞–ї–µ—З–љ—Л—Е –Њ—В–ї–Њ–ґ–µ–љ–Є–є —Г—Б—В–∞–љ–Њ–≤–ї–µ–љ—Л –µ–і–Є–љ–Є—З–љ—Л–µ —Б—В–≤–Њ—А–Ї–Є –Љ–Њ—А—Б–Ї–Є—Е –њ–ї–∞–љ–Ї—В–Њ–љ–љ—Л—Е –і–Є–∞—В–Њ–Љ–µ–є Thalassiosira gravida, –Є –Њ–±–ї–Њ–Љ–Ї–Є —Б—В–≤–Њ—А–Њ–Ї –Љ–Њ—А—Б–Ї–Є—Е –Є –±–µ–љ—В–Њ—Б–љ—Л—Е –і–Є–∞—В–Њ–Љ–µ–є, –µ–і–Є–љ–Є—З–љ–Њ –Њ–±–љ–∞—А—Г–ґ–µ–љ–∞ —Б—В–≤–Њ—А–Ї–∞ Rhabdonema cf. japonicum, –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ, –њ–µ—А–µ–Њ—В–ї–Њ–ґ–µ–љ–љ–∞—П –Є–Ј –љ–µ–Њ–≥–µ–љ–Њ–≤—Л—Е –Њ—В–ї–Њ–ґ–µ–љ–Є–є.
–Т—Л—И–µ –њ–Њ —А–∞–Ј—А–µ–Ј—Г —Н—В–Њ–є –њ–∞—З–Ї–Є –∞–ї–µ–≤—А–Є—В–Њ–≤: –≤ –Њ–±—А–∞–Ј—Ж–∞—Е 3298/2, 3298/14, 3298/16 –Є 3298/19 —Г—Б—В–∞–љ–Њ–≤–ї–µ–љ—Л –Њ–±–µ–і–љ–µ–љ–љ—Л–µ –Ї–Њ–ї–Є—З–µ—Б—В–≤–µ–љ–љ–Њ –Љ–Њ—А—Б–Ї–Є–µ –і–Є–∞—В–Њ–Љ–Њ–≤—Л–µ –Ї–Њ–Љ–њ–ї–µ–Ї—Б—Л, –≤ –Ї–Њ—В–Њ—А—Л—Е –љ–∞–Є–±–Њ–ї–µ–µ –Ј–∞–Љ–µ—В–љ—Л: –±–µ–љ—В–Њ—Б–љ—Л–µ ¬Ђ—А–µ–і–Ї–Њ¬ї Grammatophora angulosa, Diploneis smithii, D. subcincta, Cocconeis costata, C. scutellum; –њ–ї–∞–љ–Ї—В–Њ–љ–љ—Л–µ ¬Ђ–µ–і–Є–љ–Є—З–љ–Њ¬ї Chaetoceros sp., Thalassiosira gravida, T. nordenskio–µldii, Porosira glacialis –Є –і—А—Г–≥–Є–µ. –£—Б—В–∞–љ–Њ–≤–ї–µ–љ–љ—Л–µ –і–Є–∞—В–Њ–Љ–Њ–≤—Л–µ –≤–Њ–і–Њ—А–Њ—Б–ї–Є –њ–Њ–Ј–≤–Њ–ї—П—О—В —Б–і–µ–ї–∞—В—М –≤—Л–≤–Њ–і –Њ —Д–Њ—А–Љ–Є—А–Њ–≤–∞–љ–Є–Є –Њ—В–ї–Њ–ґ–µ–љ–Є–є –≤ —Б—Г–±–ї–Є—В–Њ—А–∞–ї—М–љ–Њ–є –Ј–Њ–љ–µ –Љ–Њ—А—Б–Ї–Њ–≥–Њ –±–∞—Б—Б–µ–є–љ–∞, —Б–Њ–ї–Њ–љ–Њ–≤–∞—В–Њ–≤–Њ–і–љ—Л–µ –Є —Б–Њ–ї–Њ–љ–Њ–≤–∞—В–Њ-–Љ–Њ—А—Б–Ї–Є–µ –і–Є–∞—В–Њ–Љ–µ–Є —Е–∞—А–∞–Ї—В–µ—А–Є–Ј—Г—О—В –љ–µ–Ї–Њ—В–Њ—А–Њ–µ –Њ–њ—А–µ—Б–љ–µ–љ–Є–µ —Н—В–Њ–є —Б—Г–±–ї–Є—В–Њ—А–∞–ї—М–љ–Њ–є –Ј–Њ–љ—Л.
–Ґ–∞–Ї–Є–Љ –Њ–±—А–∞–Ј–Њ–Љ, –≤ –Њ–±–љ–∞–ґ–µ–љ–Є–Є I, –≤ –љ–Є–ґ–љ–µ–є —З–∞—Б—В–Є –∞–ї–µ–≤—А–Є—В–Њ–≤–Њ–є –њ–∞—З–Ї–Є —Г—Б—В–∞–љ–Њ–≤–ї–µ–љ –Љ–Њ—А—Б–Ї–Њ–є –і–Є–∞—В–Њ–Љ–Њ–≤—Л–є –Ї–Њ–Љ–њ–ї–µ–Ї—Б, –Њ–њ—А–µ–і–µ–ї—П—О—Й–Є–є –њ—А–Є–±—А–µ–ґ–љ—Л–µ —Г—Б–ї–Њ–≤–Є—П, –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ, –±–Њ–ї–µ–µ –≥–ї—Г–±–Њ–Ї–Њ–≤–Њ–і–љ—Л–µ, –њ–Њ—Б–Ї–Њ–ї—М–Ї—Г –њ–ї–∞–љ–Ї—В–Њ–љ–љ–∞—П –≥—А—Г–њ–њ–∞ –і–Є–∞—В–Њ–Љ–µ–є –≤ –і–Є–∞—В–Њ–Љ–Њ–≤—Л—Е –Ї–Њ–Љ–њ–ї–µ–Ї—Б–∞—Е –Ј–∞–Љ–µ—В–љ–µ–µ –Є –њ–Њ –≤–Є–і–Њ–≤–Њ–Љ—Г —Б–Њ—Б—В–∞–≤—Г –Є –Ї–Њ–ї–Є—З–µ—Б—В–≤–µ–љ–љ–Њ. –Т—Л—И–µ –њ–Њ —А–∞–Ј—А–µ–Ј—Г –і–Є–∞—В–Њ–Љ–Њ–≤—Л–µ –Ї–Њ–Љ–њ–ї–µ–Ї—Б—Л –Њ—З–µ–љ—М –њ–Њ—Е–Њ–ґ–Є –њ–Њ –≤–Є–і–Њ–≤–Њ–Љ—Г —Б–Њ—Б—В–∞–≤—Г, –љ–Њ –±–µ–љ—В–Њ—Б–љ—Л–µ –≤–Є–і—Л –±–Њ–ї–µ–µ —А–∞–Ј–љ–Њ–Њ–±—А–∞–Ј–љ—Л –≤ –≤–Є–і–Њ–≤–Њ–Љ –Њ—В–љ–Њ—И–µ–љ–Є–Є –Є –±–Њ–ї–µ–µ –њ—А–µ–і—Б—В–∞–≤–Є—В–µ–ї—М–љ—Л –Ї–Њ–ї–Є—З–µ—Б—В–≤–µ–љ–љ–Њ –Є —Е–∞—А–∞–Ї—В–µ—А–Є–Ј—Г—О—В –Љ–Њ—А—Б–Ї–Є–µ —Б—Г–±–ї–Є—В–Њ—А–∞–ї—М–љ—Л–µ —Г—Б–ї–Њ–≤–Є—П.
–Т—Л—И–µ –њ–Њ —А–∞–Ј—А–µ–Ј—Г –≤ –њ–∞—З–Ї–µ –≥–ї–Є–љ –Њ–±—А–∞–Ј—Ж–∞—Е 3298/6, 3298/8, 3298/9 –Є 3298/10 —Г—Б—В–∞–љ–Њ–≤–ї–µ–љ—Л –Љ–Њ—А—Б–Ї–Є–µ –Њ–±–µ–і–љ–µ–љ–љ—Л–µ –≤ –≤–Є–і–Њ–≤–Њ–Љ –Є –Ї–Њ–ї–Є—З–µ—Б—В–≤–µ–љ–љ–Њ–Љ –Њ—В–љ–Њ—И–µ–љ–Є–Є –і–Є–∞—В–Њ–Љ–Њ–≤—Л–µ –Ї–Њ–Љ–њ–ї–µ–Ї—Б—Л, –≤ –Њ–±—А–∞–Ј—Ж–µ 3298/9 –Њ–±–љ–∞—А—Г–ґ–µ–љ–Њ –љ–∞–Є–±–Њ–ї–µ–µ –Ї–Њ–ї–Є—З–µ—Б—В–≤–Њ —Б—В–≤–Њ—А–Њ–Ї –і–Є–∞—В–Њ–Љ–µ–є –Є –Ї–Њ–ї–Є—З–µ—Б—В–≤–Њ –≤–Є–і–Њ–≤. –Т –і–Є–∞—В–Њ–Љ–Њ–≤—Л—Е –Ї–Њ–Љ–њ–ї–µ–Ї—Б–∞—Е –≤—Б—В—А–µ—З–µ–љ—Л: –љ–µ—А–Є—В–Є—З–µ—Б–Ї–Є–µ –њ–ї–∞–љ–Ї—В–Њ–љ–љ—Л–µ Thalassiosira gravida, –ї–µ–і–Њ–≤–Њ-–Љ–Њ—А—Б–Ї–Є–µ –њ–ї–∞–љ–Ї—В–Њ–љ–љ—Л–µ Fragilariopsis cylindrus, Melosira arctica, –±–µ–љ—В–Њ—Б–љ—Л–µ Cocconeis costata, C. scutellum, Diploneis smithii, Grammatophora angulosa, Navicula directa, Pseudogomphonema cf. septentrionale –Є –і—А—Г–≥–Є–µ, –Њ–±–ї–Њ–Љ–Ї–Є —Б—В–≤–Њ—А–Њ–Ї –±–µ–љ—В–Њ—Б–љ–Њ–≥–Њ –≤–Є–і–∞ Trachyneis aspera.
–Ь–Њ–ґ–љ–Њ –њ—А–µ–і–њ–Њ–ї–Њ–ґ–Є—В—М, —З—В–Њ –і–Є–∞—В–Њ–Љ–Њ–≤—Л–µ —Б—В–≤–Њ—А–Ї–Є –љ–∞–Ї–∞–њ–ї–Є–≤–∞–ї–Є—Б—М –≤ —Б—Г–±–ї–Є—В–Њ—А–∞–ї—М–љ–Њ–є –Ј–Њ–љ–µ –Љ–Њ—А—Б–Ї–Њ–≥–Њ –±–∞—Б—Б–µ–є–љ–∞.
–Т –Њ–±—А–∞–Ј—Ж–µ 3298/8 –Њ–±–љ–∞—А—Г–ґ–µ–љ –і–Є–∞—В–Њ–Љ–Њ–≤—Л–є –Ї–Њ–Љ–њ–ї–µ–Ї—Б, —Б–Њ—Б—В–Њ—П—Й–Є–є –Є–Ј –Љ–Њ—А—Б–Ї–Є—Е –і–Є–∞—В–Њ–Љ–µ–є, –њ—А–µ–і—Б—В–∞–≤–ї–µ–љ–љ—Л–є –µ–і–Є–љ–Є—З–љ—Л–Љ–Є —Ж–µ–ї—Л–Љ–Є —Б—В–≤–Њ—А–Ї–∞–Љ–Є, –≤ –±–Њ–ї—М—И–Є–љ—Б—В–≤–µ –ґ–µ –і–Є–∞—В–Њ–Љ–µ–Є –њ—А–µ–і—Б—В–∞–≤–ї–µ–љ—Л –Њ–±–ї–Њ–Љ–Ї–∞–Љ–Є, –Ї–Њ—В–Њ—А—Л–µ –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ –Є–і–µ–љ—В–Є—Д–Є—Ж–Є—А–Њ–≤–∞—В—М, –Є –Њ—З–µ–љ—М –Љ–∞–ї–µ–љ—М–Ї–Є–Љ–Є —Д—А–∞–≥–Љ–µ–љ—В–∞–Љ–Є —Б—В–≤–Њ—А–Њ–Ї. –Ь–Њ–ґ–љ–Њ –њ—А–µ–і–њ–Њ–ї–Њ–ґ–Є—В—М, —З—В–Њ –і–Є–∞—В–Њ–Љ–Њ–≤—Л–µ —Б—В–≤–Њ—А–Ї–Є –љ–∞–Ї–∞–њ–ї–Є–≤–∞–ї–Є—Б—М –≤ —Б—Г–±–ї–Є—В–Њ—А–∞–ї—М–љ–Њ–є –Ј–Њ–љ–µ –Љ–Њ—А—Б–Ї–Њ–≥–Њ –±–∞—Б—Б–µ–є–љ–∞, –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ, –≤ –µ–≥–Њ –њ—А–Є–±–Њ–є–љ–Њ–є —З–∞—Б—В–Є, —З–µ–Љ –њ—А–µ–і–њ–Њ–ї–Њ–ґ–Є—В–µ–ї—М–љ–Њ –Љ–Њ–ґ–љ–Њ –Њ–±—К—П—Б–љ–Є—В—М –Є—Е –њ–ї–Њ—Е—Г—О —Б–Њ—Е—А–∞–љ–љ–Њ—Б—В—М.
–Т –Њ–±—А–∞–Ј—Ж–µ 3298/11 —Г—Б—В–∞–љ–Њ–≤–ї–µ–љ—Л —В–Њ–ї—М–Ї–Њ –Њ–±–ї–Њ–Љ–Ї–Є —Б—В–≤–Њ—А–Њ–Ї –Љ–Њ—А—Б–Ї–Є—Е –і–Є–∞—В–Њ–Љ–µ–є.
–Я—А–Є —Б—А–∞–≤–љ–µ–љ–Є–Є –і–Є–∞—В–Њ–Љ–Њ–≤—Л—Е –≤–Њ–і–Њ—А–Њ—Б–ї–µ–є –Є–Ј –њ–∞—З–µ–Ї (–њ–∞—З–Ї–∞ I) –∞–ї–µ–≤—А–Є—В–Њ–≤ –Є –≥–ї–Є–љ (–њ–∞—З–Ї–∞ II) –Љ–Њ–ґ–љ–Њ –Ј–∞–Ї–ї—О—З–Є—В—М, —З—В–Њ –≤–µ—А—Е–љ—П—П —З–∞—Б—В—М –∞–ї–µ–≤—А–Є—В–Њ–≤ –Є –≥–ї–Є–љ—Л —Д–Њ—А–Љ–Є—А–Њ–≤–∞–ї–Є—Б—М –≤ —Б—Г–±–ї–Є—В–Њ—А–∞–ї—М–љ–Њ–є —З–∞—Б—В–Є –Љ–Њ—А—Б–Ї–Њ–≥–Њ –±–∞—Б—Б–µ–є–љ–∞, –Є —В–Њ–ї—М–Ї–Њ –љ–Є–ґ–љ—П—П —З–∞—Б—В—М –∞–ї–µ–≤—А–Є—В–Њ–≤ (–Њ–±—А–∞–Ј–µ—Ж 3298/21), –≤–µ—А–Њ—П—В–љ–Њ, —Д–Њ—А–Љ–Є—А–Њ–≤–∞–ї–∞—Б—М –≤ –±–Њ–ї–µ–µ –≥–ї—Г–±–Њ–Ї–Њ–≤–Њ–і–љ—Л—Е —Г—Б–ї–Њ–≤–Є—П—Е.
–Ґ–∞–±–ї–Є—Ж–∞ 1. –Ф–Є–∞—В–Њ–Љ–µ–Є –≤ —А–∞–Ј—А–µ–Ј–µ 3298 –∞—А—Е–Є–њ–µ–ї–∞–≥ –°–µ–≤–µ—А–љ–∞—П –Ч–µ–Љ–ї—П, –Њ—Б—В—А–Њ–≤ –С–Њ–ї—М—И–µ–≤–Є–Ї
|
–Ф–Є–∞—В–Њ–Љ–Њ–≤—Л–µ –≤–Њ–і–Њ—А–Њ—Б–ї–Є |
вДЦ –Њ–±—А–∞–Ј—Ж–Њ–≤ |
|||||||||||
|
3298-21 |
3298-20 |
3298-19 |
3298-2 |
3298-14 |
3298-16 |
3298-4 |
3298-6 |
3298-8 |
3298-9 |
3298-10 |
3298-11 |
|
|
–Ю–Ї–µ–∞–љ–Є—З–µ—Б–Ї–Є–µ –Є –љ–µ—А–Є—В–Є—З–µ—Б–Ї–Є–µ |
||||||||||||
|
A—Бtinocyclus sp. |
vr |
vr |
vr |
|||||||||
|
Bacterosira fragilis Gran |
vr |
fr. |
||||||||||
|
Chaetoceros septentrionalis √Шstrup |
vr |
|||||||||||
|
Ch. sp. |
vr |
vr |
vr |
vr |
||||||||
|
Fragilariopsis cylindrus (Grunow) Krieger |
vr |
|||||||||||
|
F. oceanica (Cleve) Hasle |
vr |
|||||||||||
|
Hyalodiscus obsoletus Sheshukova |
vr |
|||||||||||
|
Melosira arctica (Ehrenberg) Dickie |
vr |
|||||||||||
|
Porosira glacialis (Grunow) Jorgensen |
vr |
|||||||||||
|
Rhizosolenia styliformis Brighwell |
vr |
|||||||||||
|
Thalassiosira baltica (Grunow) Ostenfeld |
||||||||||||
|
T. decipiens (Grunow) Jorgensen |
vr |
|||||||||||
|
T. gravida Cleve |
f |
vr |
vr |
vr |
vr |
vr |
vr |
vr |
vr |
vr |
vr |
|
|
T. nordenskio–µldii Cleve |
vr |
|||||||||||
|
Centricae |
fr. |
fr. |
fr. |
fr. |
fr. |
fr. |
fr. |
fr. |
fr. |
fr. |
fr. |
fr. |
|
–С–µ–љ—В–Є—З–µ—Б–Ї–Є–µ –Љ–Њ—А—Б–Ї–Є–µ |
||||||||||||
|
Amphora proteus Gregory |
fr. |
fr. |
||||||||||
|
Bacillaria socialis (Gregory) Grunow |
fr. |
fr. |
fr. |
fr. |
||||||||
|
Cocconeis costata Gregory |
fr. |
fr. |
||||||||||
|
C. scutellum Ehrenberg |
r |
vr |
vr |
r |
vr |
vr |
vr |
vr |
vr |
|||
|
Diploneis interrupta (K√Љtzing) Cleve |
vr |
vr |
||||||||||
|
D. smithii (Brébisson) Cleve |
vr |
vr |
vr |
vr |
fr. |
vr |
vr |
vr |
vr |
fr. |
||
|
D. subcincta (Schmidt) Cleve |
vr |
vr |
fr. |
fr. |
vr |
fr. |
||||||
|
Fallacia pygmaea (K√Љtzing) Stickle & D.G. Mann |
vr |
|||||||||||
|
Grammatophora angulosa Ehrenberg |
vr |
f |
vr |
vr |
vr |
r |
vr |
|||||
|
Gyrosigma sp. |
fr. |
|||||||||||
|
Hannaea arcus (Ehrenberg) Patrick |
fr. |
|||||||||||
|
–Ф–Є–∞—В–Њ–Љ–Њ–≤—Л–µ –≤–Њ–і–Њ—А–Њ—Б–ї–Є |
вДЦ –Њ–±—А–∞–Ј—Ж–Њ–≤ |
|||||||||||
|
3298-21 |
3298-20 |
3298-19 |
3298-2 |
3298-14 |
3298-16 |
3298-4 |
3298-6 |
3298-8 |
3298-9 |
3298-10 |
3298-11 |
|
|
Lyrella lyra (Ehrenberg) Karaeva |
vr |
vr |
vr |
|||||||||
|
Navicula directa (Smith) Ralfs |
vr |
vr |
vr |
|||||||||
|
N. giacialis (Cleve) Grunow |
fr. |
|||||||||||
|
N. transistans Cleve |
vr |
|||||||||||
|
N. sp. |
vr |
vr |
vr |
vr |
fr. |
|||||||
|
Nitzschia sp. |
vr |
fr. |
fr. |
fr. |
fr. |
vr |
||||||
|
Pleurosigma sp. |
fr. |
|||||||||||
|
Pseudogomphonema cf. septentrionale (√Шstrup) Medlin |
vr |
vr |
||||||||||
|
Rhabdonema sp. (—Б–µ–њ—В—Л) |
vr |
vr |
fr. |
|||||||||
|
Trachyneis aspera (Ehrenberg) Cleve |
vr |
fr. |
fr. |
fr. |
fr. |
fr. |
fr. |
vr |
fr. |
|||
|
Pennatae |
fr. |
fr. |
fr. |
fr. |
fr. |
fr. |
fr. |
fr. |
fr. |
fr. |
fr. |
fr. |
|
|
||||||||||||
|
–С–µ–љ—В–Є—З–µ—Б–Ї–Є–µ –њ—А–µ—Б–љ–Њ–≤–Њ–і–љ—Л–µ |
||||||||||||
|
Amphora sp. |
vr |
|||||||||||
|
Fragilaria spp. |
r |
r |
vr |
|||||||||
|
Pinnularia obscura Krasske |
vr |
|||||||||||
|
Psammothidium sp. |
vr |
|||||||||||
|
Ulnaria ulna (Nitzsch) Compère |
fr. |
fr. |
fr. |
fr. |
||||||||
|
|
||||||||||||
|
–Ґ—А–µ—В–Є—З–љ—Л–µ –і–Є–∞—В–Њ–Љ–µ–Є |
||||||||||||
|
Rhabdonema cf. japonicum Tempère & Brun |
vr |
|||||||||||
|
|
||||||||||||
|
Chrysophycean cysts |
vr |
vr |
vr |
vr |
vr |
|||||||
–Я—А–Є–Љ–µ—З–∞–љ–Є–µ.
vr - very rare - –µ–і–Є–љ–Є—З–љ–Њ (1-5 —Б—В–≤–Њ—А–Њ–Ї –љ–∞ –њ—А–µ–њ–∞—А–∞—В)
r - rare - —А–µ–і–Ї–Њ (6-30)
f вАУ frequently - –љ–µ—А–µ–і–Ї–Њ (31-60);
fr. вАУ –Њ–±–ї–Њ–Љ–Ї–Є —Б—В–≤–Њ—А–Њ–Ї –і–Є–∞—В–Њ–Љ–µ–є
–§–Њ—А–∞–Љ–Є–љ–Є—Д–µ—А—Л –≤ –Њ—Б–∞–і–Ї–∞—Е –Њ–њ—А–µ–і–µ–ї—П–ї–∞ –Љ–Є–Ї—А–Њ–њ–∞–ї–µ–Њ–љ—В–Њ–ї–Њ–≥ –Э. –Ѓ. –Р–љ–Є–Ї–Є–љ–∞.
–Т –Њ–±—А–∞–Ј—Ж–∞—Е вДЦвДЦ 3298/1 –Є 3298/2 –Њ–њ—А–µ–і–µ–ї–µ–љ—Л –≤ —Г–Љ–µ—А–µ–љ–љ–Њ–Љ –Ї–Њ–ї–Є—З–µ—Б—В–≤–µ –∞–≥–≥–ї—О—В–Є–љ–Є—А—Г—О—Й–Є–µ —Д–Њ—А–∞–Љ–Є–љ–Є—Д–µ—А—Л Saccammina sp. –≠—В–Њ—В –≤–Є–і –Є–Љ–µ–µ—В —И–Є—А–Њ–Ї–Њ–µ –≤–Њ–Ј—А–∞—Б—В–љ–Њ–µ —А–∞—Б–њ—А–Њ—Б—В—А–∞–љ–µ–љ–Є–µ –Њ—В —Б–Є–ї—Г—А–∞ –і–Њ –љ—Л–љ–µ. –≠—В–Є —Д–Њ—А–∞–Љ–Є–љ–Є—Д–µ—А—Л ¬ЂвА¶ –њ–Њ—З—В–Є –њ–Њ–≤—Б–µ–Љ–µ—Б—В–љ–Њ —А–∞—Б–њ—А–Њ—Б—В—А–∞–љ–µ–љ—Л –≤ –Љ–µ–Ј–Њ–Ј–Њ–µ –Є –Ї–∞–є–љ–Њ–Ј–Њ–µ, —А–∞—Б—Б–µ–ї–µ–љ—Л –≤ —Б–Њ–≤—А–µ–Љ–µ–љ–љ—Л—Е –Љ–Њ—А—П—Е, –Њ–±—А–∞–Ј—Г—П –≤ —Е–Њ–ї–Њ–і–љ–Њ–≤–Њ–і–љ—Л—Е —Г—Б–ї–Њ–≤–Є—П—Е –Љ–∞—Б—Б–Њ–≤—Л–µ —Б–Ї–Њ–њ–ї–µ–љ–Є—П¬ї. [–Т–≤–µ–і–µ–љ–Є–µ –≤ –Є–Ј—Г—З–µ–љ–Є–µ —Д–Њ—А–∞–Љ–Є–љ–Є—Д–µ—А. –Я–Њ–і —А–µ–і–∞–Ї—Ж–Є–µ–є –Э. –Э. –°—Г–±–±–Њ—В–Є–љ–Њ–є, –Э. –Р. –Т–Њ–ї–Њ—И–Є–љ–Њ–≤–Њ–є, –Р.–ѓ. –Р–Ј–±–µ–ї—М, 1981–≥.].
–Ю–±—А–∞–Ј–µ—Ж вДЦ3298/6 —Б–Њ–і–µ—А–ґ–Є—В –і–Њ—Б—В–∞—В–Њ—З–љ–Њ –Љ–љ–Њ–≥–Њ—З–Є—Б–ї–µ–љ–љ—Л–є –∞—А–Ї—В–Њ-–±–Њ—А–µ–∞–ї—М–љ—Л–є –Ї–Њ–Љ–њ–ї–µ–Ї—Б, –≤ –Ї–Њ—В–Њ—А–Њ–Љ 26 –≤–Є–і–Њ–≤ –Є 796 —Н–Ї–Ј–µ–Љ–њ–ї—П—А–Њ–≤ —А–∞–Ї–Њ–≤–Є–љ–Њ–Ї —Д–Њ—А–∞–Љ–Є–љ–Є—Д–µ—А –≤ –Њ—Б–љ–Њ–≤–љ–Њ–Љ —Е–Њ—А–Њ—И–µ–є –Є —Б—А–µ–і–љ–µ–є —Б–Њ—Е—А–∞–љ–љ–Њ—Б—В–Є.
–Т —Н—В–Њ–Љ –Њ–±—А–∞–Ј—Ж–µ —П–≤–љ–Њ–µ –њ—А–µ–Њ–±–ї–∞–і–∞–љ–Є–µ –Ї–∞—Б—Б–Є–і—Г–ї–Є–љ–Є–і, —Б—А–µ–і–Є –Ї–Њ—В–Њ—А—Л—Е –њ—А–µ–Њ–±–ї–∞–і–∞–µ—В –≤–Є–і Cassandra teretis —Б —Е–Њ—А–Њ—И–Њ —А–∞–Ј–≤–Є—В—Л–Љ–Є –Ї—А—Г–њ–љ—Л–Љ–Є —А–∞–Ї–Њ–≤–Є–љ–Ї–∞–Љ–Є. –Ю–і–љ–∞–Ї–Њ –њ–Њ–њ–∞–і–∞—О—В—Б—П —Б—А–µ–і–Є –љ–Є—Е –Љ–µ–ї–Ї–Є–µ, –і–µ—Д–Њ—А–Љ–Є—А–Њ–≤–∞–љ–љ—Л–µ –Є —Г—А–Њ–і–ї–Є–≤—Л–µ (—Б—А–Њ—Б—И–Є–µ—Б—П) –Њ—Б–Њ–±–Є. –≠—В–Њ—В –∞—А–Ї—В–Є—З–µ—Б–Ї–Є–є –≤–Є–і –Њ–±–Є—В–∞–µ—В –≤ –њ—А–µ–і–µ–ї–∞—Е –Љ–∞—В–µ—А–Є–Ї–Њ–≤–Њ–є –Њ—В–Љ–µ–ї–Є. –Ь–µ–љ—М—И–µ —А–∞–Ї–Њ–≤–Є–љ–Њ–Ї Cassidulina reniforme –Є C. subacuta.
–†–µ—В—А–Њ—Н–ї—М—Д–Є–і–Є—Г–Љ—Л –Љ–∞–ї–Њ—З–Є—Б–ї–µ–љ–љ—Л –Є –њ—А–µ–і—Б—В–∞–≤–ї–µ–љ—Л –Љ–µ–ї–Ї–Є–Љ–Є, —З–∞—Б—В–Њ –ї–Њ–Љ–∞–љ—Л–Љ–Є —А–∞–Ї–Њ–≤–Є–љ–Ї–∞–Љ–Є –Њ–і–љ–Њ–≥–Њ –≤–Є–і–∞ Retroelphidium atlanticum.
–Э–Њ–љ–Є–Њ–љ–Є–і—Л Cribrononion obscurus, Cr.incertus, Nonionelina labradorica –Є Melonis zaandamae –µ–і–Є–љ–Є—З–љ—Л, —В–Њ–ї—М–Ї–Њ –≤–Є–і Astrononion gallowayi –і–Њ—Б—В–∞—В–Њ—З–љ–Њ –Љ–љ–Њ–≥–Њ—З–Є—Б–ї–µ–љ–µ–љ (74 —Н–Ї–Ј–µ–Љ–њ–ї—П—А–∞) –Є –Є–Љ–µ–µ—В –Љ–µ–ї–Ї–Є–µ, –≤ –Њ—Б–љ–Њ–≤–љ–Њ–Љ –ї–Њ–Љ–∞–љ—Л–µ —А–∞–Ї–Њ–≤–Є–љ–Ї–Є. –Т—Л–і–µ–ї–µ–љ—Л —В–∞–Ї–ґ–µ —Е–∞–є–љ–µ–Ј–Є–љ—Л –≤–Є–і–∞ Haynesina orbiculare. –Ш–Ј –Ї—А–Є–±—А–Њ—Н–ї—М—Д–Є–і–Є—Г–Љ–Њ–≤ –Њ–њ—А–µ–і–µ–ї–µ–љ—Л Cribroelphidium goesi. –Я—А–Є—Б—Г—В—Б—В–≤—Г—О—В –±—Г–Ї—Ж–µ–ї–ї—Л –≤–Є–і–Њ–≤ Buccella frigida –Є B. hannai arctica, –Њ–њ—А–µ–і–µ–ї–µ–љ—Л –µ–і–Є–љ–Є—З–љ—Л–µ Pseudopolimorphina novangliae, Hyalinea sp., Asterigerinata pulchella, Alabaminoides mitis, Gavelinopsis praegeri, Bolivina pseudopunctata, Fursenkoina gracilis –Є –і—А—Г–≥–Є–µ.
–Х–і–Є–љ—Б—В–≤–µ–љ–љ–∞—П Quinqueloculina sp. –Є–Љ–µ–µ—В –ї–Њ–Љ–∞–љ—Г—О, –њ–Њ—В—С—А—В—Г—О —А–∞–Ї–Њ–≤–Є–љ–Ї—Г –Є —П–≤–ї—П–µ—В—Б—П, –≤–µ—А–Њ—П—В–љ–µ–є –≤—Б–µ–≥–Њ, –њ–µ—А–µ–Њ—В–ї–Њ–ґ–µ–љ–љ–Њ–є.
–Я—А–Є—Б—Г—В—Б—В–≤—Г—О—Й–Є–є –≤ —Н—В–Њ–Љ –Ї–Њ–Љ–њ–ї–µ–Ї—Б–µ –≤–Є–і Cassidulina reniforme —П–≤–ї—П–µ—В—Б—П –Љ–µ–ї–Ї–Њ–≤–Њ–і–љ–Њ-–Љ–Њ—А—Б–Ї–Є–Љ, –∞ –µ–≥–Њ –і–Њ—Б—В–∞—В–Њ—З–љ–Њ –±–Њ–ї—М—И–Њ–µ –Ї–Њ–ї–Є—З–µ—Б—В–≤–Њ —Б–≤–Є–і–µ—В–µ–ї—М—Б—В–≤—Г–µ—В –Њ –љ–µ–±–ї–∞–≥–Њ–њ—А–Є—П—В–љ—Л—Е —Г—Б–ї–Њ–≤–Є—П—Е –і–ї—П –ґ–Є–Ј–љ–Є –і—А—Г–≥–Є—Е —Д–Њ—А–∞–Љ–Є–љ–Є—Д–µ—А. –≠—В–Њ –њ–Њ–і—В–≤–µ—А–ґ–і–∞–µ—В—Б—П –Є –Љ–µ–ї–Ї–Є–Љ–Є —А–∞–Ј–Љ–µ—А–∞–Љ–Є –±–Њ–ї—М—И–Є–љ—Б—В–≤–∞ –Њ—Б—В–∞–ї—М–љ—Л—Е —А–∞–Ї–Њ–≤–Є–љ–Њ–Ї –Ї–Њ–Љ–њ–ї–µ–Ї—Б–∞, –∞ —В–∞–Ї–ґ–µ –њ—А–Є—Б—Г—В—Б—В–≤–Є–µ–Љ —Г—А–Њ–і–ї–Є–≤—Л—Е —Д–Њ—А–Љ.
–Ш—Б—Е–Њ–і—П –Є–Ј –њ–Њ–ї—Г—З–µ–љ–љ—Л—Е –і–∞–љ–љ—Л—Е, –Љ–Њ–ґ–љ–Њ –њ—А–µ–і–њ–Њ–ї–Њ–ґ–Є—В—М, —З—В–Њ –∞—А–Ї—В–Њ-–±–Њ—А–µ–∞–ї—М–љ—Л–є –Ї–Њ–Љ–њ–ї–µ–Ї—Б —Н—В–Њ–≥–Њ –Њ–±—А–∞–Ј—Ж–∞ —Д–Њ—А–Љ–Є—А–Њ–≤–∞–ї—Б—П –≤ –Њ—В–Ї—А—Л—В–Њ–Љ –Њ—В–љ–Њ—Б–Є—В–µ–ї—М–љ–Њ –љ–µ–≥–ї—Г–±–Њ–Ї–Њ–Љ –Љ–Њ—А—Б–Ї–Њ–Љ –±–∞—Б—Б–µ–є–љ–µ –≤ –њ—А–µ–і–µ–ї–∞—Е —И–µ–ї—М—Д–∞, c –љ–Є–Ј–Ї–Њ–є –њ—А–Є–і–Њ–љ–љ–Њ–є —В–µ–Љ–њ–µ—А–∞—В—Г—А–Њ–є –≤–Њ–і –Є —Б —Б–Њ–ї—С–љ–Њ—Б—В—М—О –±–ї–Є–Ј–Ї–Њ–є –Ї –љ–Њ—А–Љ–∞–ї—М–љ–Њ-–Љ–Њ—А—Б–Ї–Њ–є.
–Ю–±—А–∞–Ј–µ—Ж вДЦ 3298/8 —Б–Њ–і–µ—А–ґ–Є—В –љ–µ–Љ–љ–Њ–≥–Њ—З–Є—Б–ї–µ–љ–љ—Л–є –∞—А–Ї—В–Њ-–±–Њ—А–µ–∞–ї—М–љ—Л–є –Ї–Њ–Љ–њ–ї–µ–Ї—Б, –≤ –Ї–Њ—В–Њ—А–Њ–Љ –њ—А–Є—Б—Г—В—Б—В–≤—Г—О—В 15–≤–Є–і–Њ–≤ –Є 58 —Н–Ї–Ј–µ–Љ–њ–ї—П—А–Њ–≤ —Д–Њ—А–∞–Љ–Є–љ–Є—Д–µ—А –≤ –Њ—Б–љ–Њ–≤–љ–Њ–Љ —Е–Њ—А–Њ—И–µ–є —Б–Њ—Е—А–∞–љ–љ–Њ—Б—В–Є.
–Т –Ї–Њ–Љ–њ–ї–µ–Ї—Б–µ –љ–∞–±–ї—О–і–∞–µ—В—Б—П —П–≤–љ–Њ–µ –њ—А–µ–Њ–±–ї–∞–і–∞–љ–Є–µ –Ї–∞—Б—Б–Є–і—Г–ї–Є–љ–Є–і –≤–Є–і–Њ–≤ Cassidulina reniforme, Cassidulina subacuta, Cassandra teretis. –°—А–µ–і–Є –њ–Њ—Б–ї–µ–і–љ–Є—Е –њ—А–µ–Њ–±–ї–∞–і–∞—О—В –Ї—А—Г–њ–љ—Л–µ, —Е–Њ—А–Њ—И–Њ —Б—Д–Њ—А–Љ–Є—А–Њ–≤–∞–љ–љ—Л–µ –Њ—Б–Њ–±–Є. –Т–Є–і Cassandra teretis —А–∞—Б–њ—А–Њ—Б—В—А–∞–љ—С–љ –≤ –∞—А–Ї—В–Є—З–µ—Б–Ї–Њ–є –Є –±–Њ—А–µ–∞–ї—М–љ–Њ–є –Њ–±–ї–∞—Б—В—П—Е, –љ–Њ —В—П–≥–Њ—В–µ–µ—В –Ї –±–Њ–ї–µ–µ —Е–Њ–ї–Њ–і–љ—Л–Љ –≤–Њ–і–љ—Л–Љ –Љ–∞—Б—Б–∞–Љ –≤ —И–µ–ї—М—Д–Њ–≤—Л—Е –Љ–Њ—А—П—Е.
–Ф–Њ—Б—В–∞—В–Њ—З–љ–Њ –Љ–љ–Њ–≥–Њ—З–Є—Б–ї–µ–љ–љ—Л –±—Г–Ї—Ж–µ–ї–ї—Л –≤–Є–і–Њ–≤ Buccella frigida –Є Buccella hannai arctica, –Љ–µ–љ—М—И–µ Astrononion gallowayi, Alabaminoides mitis. –Х–і–Є–љ–Є—З–љ–Њ –њ—А–Є—Б—Г—В—Б—В–≤—Г—О—В Sigmomorphina undulosa, Discorbis deplanatus, Melonis sp. - juv., Haynesina orbiculare, Cribroelphidium subarcticum.
–£—З–Є—В—Л–≤–∞—П –њ–Њ–ї—Г—З–µ–љ–љ—Л–µ —А–µ–Ј—Г–ї—М—В–∞—В—Л, –Љ–Њ–ґ–љ–Њ –њ—А–µ–і–њ–Њ–ї–Њ–ґ–Є—В—М, —З—В–Њ —Н—В–Њ—В –љ–µ—Б–Ї–Њ–ї—М–Ї–Њ –Њ–±–µ–і–љ—С–љ–љ—Л–є –Ї–Њ–Љ–њ–ї–µ–Ї—Б —Д–Њ—А–Љ–Є—А–Њ–≤–∞–ї—Б—П, –≤–µ—А–Њ—П—В–љ–µ–є –≤—Б–µ–≥–Њ, –≤ –љ–µ –≥–ї—Г–±–Њ–Ї–Њ–Љ –Љ–Њ—А—Б–Ї–Њ–Љ –±–∞—Б—Б–µ–є–љ–µ —Б –љ–Њ—А–Љ–∞–ї—М–љ–Њ–є —Б–Њ–ї—С–љ–Њ—Б—В—М—О –Є –Њ—В—А–Є—Ж–∞—В–µ–ї—М–љ—Л–Љ–Є –њ—А–Є–і–Њ–љ–љ—Л–Љ–Є —В–µ–Љ–њ–µ—А–∞—В—Г—А–∞–Љ–Є –≤–Њ–і.
–Т –Њ–±—А–∞–Ј—Ж–µ вДЦ 3298/8 –Њ–±–љ–∞—А—Г–ґ–µ–љ–∞ –µ–і–Є–љ—Б—В–≤–µ–љ–љ–∞—П —А–∞–Ї–Њ–≤–Є–љ–Ї–∞ –Њ—Б—В—А–∞–Ї–Њ–і—Л
–Т –Њ–±—А–∞–Ј—Ж–∞—Е вДЦвДЦ 3298/9 –Є 3298/10 –њ—А–Є—Б—Г—В—Б—В–≤—Г—О—В –≤ –љ–µ–±–Њ–ї—М—И–Њ–Љ –Ї–Њ–ї–Є—З–µ—Б—В–≤–µ –∞–≥–≥–ї—О—В–Є–љ–Є—А—Г—О—Й–Є–µ —Д–Њ—А–∞–Љ–Є–љ–Є—Д–µ—А—Л Rhabdammina sp. –Є –Њ–±–ї–Њ–Љ–Ї–Є Textularia sp. –≠—В–Є –Љ–Њ—А—Б–Ї–Є–µ –≥–ї—Г–±–Њ–Ї–Њ–≤–Њ–і–љ—Л–µ –≤–Є–і—Л —А–∞—Б–њ—А–Њ—Б—В—А–∞–љ–µ–љ—Л –њ–Њ–≤—Б–µ–Љ–µ—Б—В–љ–Њ –Є –Є–Љ–µ—О—В —И–Є—А–Њ–Ї–Њ–µ –≤–Њ–Ј—А–∞—Б—В–љ–Њ–µ —А–∞—Б–њ—А–Њ—Б—В—А–∞–љ–µ–љ–Є–µ –Њ—В –Њ—А–і–Њ–≤–Є–Ї–∞ –і–Њ –љ—Л–љ–µ.
–Ю–±—А–∞–Ј—Ж—Л вДЦвДЦ 3298/14, 3298/20, 3298/21 —Б–Њ–і–µ—А–ґ–∞—В –µ–і–Є–љ–Є—З–љ—Л–µ —А–∞–Ї–Њ–≤–Є–љ–Ї–Є —Д–Њ—А–∞–Љ–Є–љ–Є—Д–µ—А (–Њ—В 2 –і–Њ 4 —Н–Ї–Ј–µ–Љ–њ–ї—П—А–Њ–≤, —Б–Љ. —В–∞–±–ї–Є—Ж—Г) –≤ –Њ—Б–љ–Њ–≤–љ–Њ–Љ —Е–Њ—А–Њ—И–µ–є —Б–Њ—Е—А–∞–љ–љ–Њ—Б—В–Є. –≠—В–Њ –∞–≥–≥–ї—О—В–Є–љ–Є—А—Г—О—Й–∞—П Rotaliammina orchaceae –Є —Б–µ–Ї—А–µ—Ж–Є–Њ–љ–љ—Л–µ –±–µ–љ—В–Њ—Б–љ—Л–µ Buccella frigida, Retroelphidium atlanticum, Haynesina orbiculare, Cassandra teretis. –≠—В–Є –≤–Є–і—Л –Є–Љ–µ—О—В —И–Є—А–Њ–Ї–Њ–µ –≤–Њ–Ј—А–∞—Б—В–љ–Њ–µ —А–∞—Б–њ—А–Њ—Б—В—А–∞–љ–µ–љ–Є–µ –≤ –Ї–∞–є–љ–Њ–Ј–Њ–µ –Є –Љ–Њ–ґ–љ–Њ –њ—А–µ–і–њ–Њ–ї–Њ–ґ–Є—В—М, —З—В–Њ —Б–Њ—Б—В–∞–≤ –Є —Б–Њ—Е—А–∞–љ–љ–Њ—Б—В—М –і–∞–љ–љ–Њ–≥–Њ –љ–∞–±–Њ—А–∞ —Д–Њ—А–∞–Љ–Є–љ–Є—Д–µ—А –њ—А–µ–і–њ–Њ–ї–∞–≥–∞–µ—В –љ–Њ—А–Љ–∞–ї—М–љ–Њ-–Љ–Њ—А—Б–Ї–Є–µ —Г—Б–ї–Њ–≤–Є—П (–∞—А–Ї—В–Є—З–µ—Б–Ї–Њ–µ –Љ–Њ—А–µ) –і–ї—П —Б–≤–Њ–µ–≥–Њ –Њ–±–Є—В–∞–љ–Є—П.
–Ъ–∞–є–љ–Њ–Ј–Њ–є—Б–Ї–Є–µ –Љ–Њ–ї–ї—О—Б–Ї–Є –≤ –Њ—В–ї–Њ–ґ–µ–љ–Є—П—Е –Њ–њ—А–µ–і–µ–ї–Є–ї –њ–∞–ї–µ–Њ–љ—В–Њ–ї–Њ–≥ –Р. –Т. –Ъ—А—Л–ї–Њ–≤.
–Ю–±—А–∞–Ј–µ—Ж вДЦ 3298/7
- Mya truncata Linnaeus, 1758 вАУ 18 –њ–Њ–≤—А–µ–ґ–і–µ–љ–љ—Л—Е —А–∞–Ї–Њ–≤–Є–љ, 5 –њ–Њ–≤—А–µ–ґ–і–µ–љ–љ—Л—Е —А–∞–Ї–Њ–≤–Є–љ —Б –і–≤—Г–Љ—П —Б–Њ–Љ–Ї–љ—Г—В—Л–Љ–Є —Б—В–≤–Њ—А–Ї–∞–Љ–Є (—З–∞—Б—В–Є—З–љ–Њ –њ–Њ–≤—А–µ–ґ–і–µ–љ—Л –њ—А–Є –Њ—В–±–Њ—А–µ –Є –Њ—В–Љ—Л–≤–Ї–µ) —З—В–Њ –Љ–Њ–ґ–µ—В —Б–≤–Є–і–µ—В–µ–ї—М—Б—В–≤–Њ–≤–∞—В—М –Њ–± –Є—Е –Ї–Њ—А–µ–љ–љ–Њ–Љ –Ј–∞–ї–µ–≥–∞–љ–Є–Є;
- Mya - 94 –Њ–±–ї–Њ–Љ–Ї–∞ —А–∞–Ї–Њ–≤–Є–љ;
- Hiatella arctic–∞ (Linnaeus, 1758) вАУ 1 —Ж–µ–ї–∞—П —А–∞–Ї–Њ–≤–Є–љ–∞ (—А–∞–Ј—А–Њ–Ј–љ–µ–љ–љ–∞—П —Б—В–≤–Њ—А–Ї–∞) –Є 2 –њ–Њ–≤—А–µ–ґ–і–µ–љ–љ—Л–µ —А–∞–Ї–Њ–≤–Є–љ—Л.
Mya truncata Linnaeus, 1758 - –Є–Ј–≤–µ—Б—В–µ–љ —Б –Љ–Є–Њ—Ж–µ–љ–∞, —И–Є—А–Њ–Ї–Њ —А–∞—Б–њ—А–Њ—Б—В—А–∞–љ–µ–љ–љ—Л–є –±–Њ—А–µ–∞–ї—М–љ–Њ-–∞—А–Ї—В–Є—З–µ—Б–Ї–Є–є —Ж–Є—А–Ї—Г–Љ–њ–Њ–ї—П—А–љ—Л–є –≤–Є–і (–Я–µ—В—А–Њ–≤, 1966, –Э–∞—Г–Љ–Њ–≤, 2006, McNeil, 1965). –Т—Б—В—А–µ—З–µ–љ –≤–Њ –≤—Б–µ—Е —Б–µ–≤–µ—А–љ—Л—Е –Љ–Њ—А—П—Е –†–Њ—Б—Б–Є–Є, –Љ–Њ—А–µ –С–Њ—Д–Њ—А—В–∞, —Г –±–µ—А–µ–≥–Њ–≤ –Ъ–∞–љ–∞–і—Б–Ї–Њ–≥–Њ –Р—А–Ї—В–Є—З–µ—Б–Ї–Њ–≥–Њ –∞—А—Е–Є–њ–µ–ї–∞–≥–∞, –У—А–µ–љ–ї–∞–љ–і–Є–Є, –Ш—Б–ї–∞–љ–і–Є–Є, –®–њ–Є—Ж–±–µ—А–≥–µ–љ–∞ –Є –Ч–µ–Љ–ї–Є –§—А–∞–љ—Ж–∞-–Ш–Њ—Б–Є—Д–∞. –Т –Р—В–ї–∞–љ—В–Є—З–µ—Б–Ї–Њ–Љ –Њ–Ї–µ–∞–љ–µ —А–∞—Б—Б–µ–ї—П–µ—В—Б—П –љ–∞ —О–≥ –і–Њ –С–Є—Б–Ї–∞–є—Б–Ї–Њ–≥–Њ –Ј–∞–ї–Є–≤–∞ –Є –њ-–Њ–≤–∞ –Ъ–µ–є–њ-–Ъ–Њ–і, –≤ –Ґ–Є—Е–Њ–Љ –Њ–Ї–µ–∞–љ–µ вАУ –і–Њ –Ц–µ–ї—В–Њ–≥–Њ –Љ–Њ—А—П –Є –Ј–∞–ї–Є–≤–∞ –Я—М—О–і–ґ–µ—В-–°–∞—Г–љ–і. –°—В–µ–љ–Њ—В–µ—А–Љ–љ—Л–є, —Г–Љ–µ—А–µ–љ–љ–Њ —В–µ–њ–ї–Њ–≤–Њ–і–љ—Л–є, –Њ—В–љ–Њ—Б–Є—В–µ–ї—М–љ–Њ —Б—В–µ–љ–Њ–±–∞—В–љ—Л–є, –Њ—В–љ–Њ—Б–Є—В–µ–ї—М–љ–Њ —Н–≤—А–Є—В–Њ–њ–љ—Л–є, –њ—А–µ–і–њ–Њ—З–Є—В–∞—О—Й–Є–є –Є–ї–Є—Б—В—Л–µ –Є –њ–µ—Б—З–∞–љ—Л–µ –≥—А—Г–љ—В—Л –≤–Є–і. –Т–њ–µ—А–≤—Л–µ –≤—Б—В—А–µ—З–∞–µ—В—Б—П –≤ –Љ–Є–Њ—Ж–µ–љ–µ –°–µ–≤–µ—А–љ–Њ–є –Я–∞—Ж–Є—Д–Є–Ї–Є (McNeil, 1965), –≤—Б—В—А–µ—З–∞–µ—В—Б—П –≤ –њ–ї–Є–Њ—Ж–µ–љ–µ –Ш—В–∞–ї–Є–Є –Є –С–µ–ї—М–≥–Є–Є (Marquet, 2005). –Ю—В–Љ–µ—З–µ–љ –љ–∞ –≥–ї—Г–±–Є–љ–∞—Е 5вАУ30 –Є 80вАУ180 –Љ (—А–µ–ґ–µ 0вАУ5 –Љ, 30вАУ80 –Љ –Є 180вАУ625 –Љ) –њ—А–Є —В–µ–Љ–њ–µ—А–∞—В—Г—А–µ –Њ—В - 2,0 –і–Њ 13,7¬∞–° –Є —Б–Њ–ї–µ–љ–Њ—Б—В–Є 23,9 - 35,2вА∞, –≤ –Њ—Б–љ–Њ–≤–љ–Њ–Љ –љ–∞ –Є–ї–Є—Б—В—Л—Е (34% –љ–∞—Е–Њ–і–Њ–Ї) –Є —Б–Љ–µ—И–∞–љ–љ—Л—Е (40% –љ–∞—Е–Њ–і–Њ–Ї) –≥—А—Г–љ—В–∞—Е. –Ь–Є–Њ—Ж–µ–љ-–≥–Њ–ї–Њ—Ж–µ–љ (–≤ –Р—А–Ї—В–Є–Ї–µ —А–∞–љ–љ–Є–є –њ–ї–µ–є—Б—В–Њ—Ж–µ–љ-–≥–Њ–ї–Њ—Ж–µ–љ, –њ—А–µ–Є–Љ—Г—Й–µ—Б—В–≤–µ–љ–љ–Њ –љ–µ–Њ–њ–ї–µ–є—Б—В–Њ—Ж–µ–љ (–Ь–µ—А–Ї–ї–Є–љ, –Ч–∞—А—Е–Є–і–Ј–µ, –Ш–ї—М–Є–љ–∞, 1979, –Э–∞—Г–Љ–Њ–≤, 2006 –Є –і—А.) –•–∞—А–∞–Ї—В–µ—А–Є–Ј—Г–µ—В –Љ—П–≥–Ї–Є–µ –≥–ї–Є–љ–Є—Б—В—Л–µ –Є–ї–Є –њ–µ—Б—З–∞–љ–Њ-–≥–ї–Є–љ–Є—Б—В—Л–µ –≥—А—Г–љ—В—Л, –≥–ї—Г–±–Є–љ—Л –∞—А–Ї—В–Є—З–µ—Б–Ї–Є–є-–±–Њ—А–µ–∞–ї—М–љ—Л–є –Ї–ї–Є–Љ–∞—В. –Р–±—Б–Њ–ї—О—В–љ–Њ–µ –і–∞—В–Є—А–Њ–≤–∞–љ–Є–µ –Љ–Є–є (–Ї–∞–Ї –Є —Ж–Є—А—В–Њ–і–∞—А–Є–є –Є –Є–љ—Л—Е –Ј–∞—А—Л–≤–∞—О—Й–Є—Е—Б—П –Љ–Њ–ї–ї—О—Б–Ї–Њ–≤) –љ–µ –≤—Б–µ–≥–і–∞ —Г–і–∞—З–љ–Њ –Њ—В—А–∞–ґ–∞–µ—В –≤–Њ–Ј—А–∞—Б—В –≤–Љ–µ—Й–∞—О—Й–Є—Е –Њ—В–ї–Њ–ґ–µ–љ–Є–є –Є–Ј-–Ј–∞ —Е–Є–Љ–Є—З–µ—Б–Ї–Њ–≥–Њ –Ј–∞—Б–Њ—А–µ–љ–Є—П —А–∞–Ї–Њ–≤–Є–љ, –Њ–і–љ–∞–Ї–Њ, –≤ —А—П–і–µ —Б–ї—Г—З–∞–µ–≤ (–Њ—В–Ї—А—Л—В–Њ–≥–Њ –Љ–Њ—А—П, —Б—В–∞–±–Є–ї—М–љ–Њ–≥–Њ —А–µ–ґ–Є–Љ–∞ —Б–Њ–ї–µ–љ–Њ—Б—В–Є –Є —Г–і–∞–ї–µ–љ–љ–Њ—Б—В–Є –Њ—В –±–µ—А–µ–≥–∞ –Є —Е–Њ—А–Њ—И–µ–є —Б–Њ—Е—А–∞–љ–љ–Њ—Б—В–Є —Б–∞–Љ–Є—Е —А–∞–Ї–Њ–≤–Є–љ, –Ј–∞—Е–Њ—А–Њ–љ–µ–љ–љ—Л—Е –љ–µ–њ–Њ–і–∞–ї–µ–Ї—Г –Њ—В –Љ–µ—Б—В–∞ —Б–≤–Њ–µ–≥–Њ –Њ–±–Є—В–∞–љ–Є—П) –Љ–Њ–ґ–µ—В –±—Л—В—М —Г—Б–њ–µ—И–љ—Л–Љ.
Hiatella arctica (Linnaeus, 1767) –Є–Ј–≤–µ—Б—В–љ–∞ —Б –Њ–ї–Є–≥–Њ—Ж–µ–љ–∞ (–Я–µ—В—А–Њ–≤, 1966, –Ч–µ–љ–Ї–µ–≤–Є—З, 1977, –Э–∞—Г–Љ–Њ–≤, 2006 –Є –і—А.), –≤ –Р—А–Ї—В–Є–Ї–µ - —Б –Љ–Є–Њ—Ж–µ–љ–∞. –Т –Љ–Њ—А—П—Е –∞—А–Ї—В–Є—З–µ—Б–Ї–Њ–≥–Њ –±–∞—Б—Б–µ–є–љ–∞ –Њ—В–Љ–µ—З–µ–љ –љ–∞ –≥–ї—Г–±–Є–љ–∞—Е 5вАУ30 –Є 80вАУ220 –Љ (—А–µ–ґ–µ 0вАУ5 –Љ, 30вАУ80 –Љ –Є 220вАУ1290 –Љ) –њ—А–Є —В–µ–Љ–њ–µ—А–∞—В—Г—А–µ –Њ—В - 2,0 –і–Њ 13,7¬∞–° –Є —Б–Њ–ї–µ–љ–Њ—Б—В–Є 23,9 - 35,5вА∞, –≤ –Њ—Б–љ–Њ–≤–љ–Њ–Љ –љ–∞ –Є–ї–Є—Б—В—Л—Е (34% –љ–∞—Е–Њ–і–Њ–Ї) –Є —Б–Љ–µ—И–∞–љ–љ—Л—Е (40% –љ–∞—Е–Њ–і–Њ–Ї) –≥—А—Г–љ—В–∞—Е. –Э–∞ –≥–ї—Г–±–Є–љ–∞—Е –Њ—В 30 –і–Њ 80 –Љ вАУ –њ—А–µ–Њ–±–ї–∞–і–∞–µ—В Hiatella pholadis (Linnaeus, 1771). –®–Є—А–Њ–Ї–Њ —А–∞—Б–њ—А–Њ—Б—В—А–∞–љ–µ–љ–љ—Л–є —Б—Г–±—В—А–Њ–њ–Є—З–µ—Б–Ї–Њ-–∞—А–Ї—В–Є—З–µ—Б–Ї–Є–є —Ж–Є—А–Ї—Г–Љ–њ–Њ–ї—П—А–љ—Л–є –≤–Є–і –Њ—В–Љ–µ—З–µ–љ –≤–Њ –≤—Б–µ—Е –Љ–Њ—А—П—Е –°–µ–≤–µ—А–љ–Њ–≥–Њ –Ы–µ–і–Њ–≤–Є—В–Њ–≥–Њ –Њ–Ї–µ–∞–љ–∞. –Ь–∞—Б—Б–Њ–≤—Л–µ –љ–∞—Е–Њ–і–Ї–Є —Н—В–Њ–≥–Њ –≤–Є–і–∞ —Б —В–Њ–ї—Б—В—Л–Љ–Є —А–∞–Ї–Њ–≤–Є–љ–∞–Љ–Є –њ–Њ—З—В–Є –њ—А—П–Љ–Њ—Г–≥–Њ–ї—М–љ—Л—Е –Њ—З–µ—А—В–∞–љ–Є–є –≤ –Љ–Њ–љ–Њ–≤–Є–і–Њ–≤—Л—Е –Є –Њ–±–µ–і–µ–љ–µ–љ–љ—Л—Е –Ї–Њ–Љ–њ–ї–µ–Ї—Б–∞—Е —З–∞—Б—В–Њ –≥–Њ–≤–Њ—А—П—В –Њ —Е–Њ–ї–Њ–і–љ–Њ–≤–Њ–і–љ—Л—Е —Г—Б–ї–Њ–≤–Є—П—Е –Њ–±–Є—В–∞–љ–Є—П, —З–∞—Б—В–Њ —А—П–і–Њ–Љ —Б –њ—А–Є–і–Њ–љ–љ–Њ–є –Њ–±–ї–∞—Б—В—М—О –ї–µ–і–љ–Є–Ї–Њ–≤, —Б–њ—Г—Б–Ї–∞—О—Й–Є—Е—Б—П –≤ –Љ–Њ—А–µ (–≤ –Њ—В–ї–Њ–ґ–µ–љ–Є—П—Е –њ–ї–µ–є—Б—В–Њ—Ж–µ–љ–∞ —Д–Є–Њ—А–і–∞ –£–і–і–µ–≤–∞–ї–ї–∞, –Ѓ–ґ–љ–∞—П –®–≤–µ—Ж–Є—П), –њ-–Њ–≤ –Ы–Є—В–Ї–µ (–°–µ–≤–µ—А–љ—Л–є –Њ—Б—В—А–Њ–≤, –∞—А—Е. –Э–Њ–≤–∞—П –Ч–µ–Љ–ї—П), –≤ –±–Њ—А–µ–∞–ї—М–љ—Л—Е –Ї–Њ–Љ–њ–ї–µ–Ї—Б–∞—Е —Н—В–Њ –≤–Є–і —В–∞–Ї–ґ–µ –≤—Б—В—А–µ—З–∞–µ—В—Б—П, –љ–Њ –Њ–±—Л—З–љ–Њ –љ–µ –і–Њ–Љ–Є–љ–Є—А—Г–µ—В –Є —Е–∞—А–∞–Ї—В–µ—А–Є–Ј—Г–µ—В –±–Њ–ї–µ–µ —В–Њ–љ–Ї–Њ—Б—В–µ–љ–љ—Л–Љ–Є —А–∞–Ї–Њ–≤–Є–љ–∞–Љ–Є –Њ–Ї—А—Г–≥–ї—Л—Е –Њ—З–µ—А—В–∞–љ–Є–є. –†–∞–Ї–Њ–≤–Є–љ—Л —Е–Є–∞—В–µ–ї–ї —П–≤–ї—П—О—В—Б—П –Њ–і–љ–Є–Љ –Є–Ј –ї—Г—З—И–Є—Е —А–∞–Ї–Њ–≤–Є–љ –Љ–Њ—А—Б–Ї–Є—Е –Љ–Њ–ї–ї—О—Б–Ї–Њ–≤ –і–ї—П –∞–±—Б–Њ–ї—О—В–љ–Њ–≥–Њ –і–∞—В–Є—А–Њ–≤–∞–љ–Є—П.
–°–Њ—Е—А–∞–љ–љ–Њ—Б—В—М. –†–∞–Ї–Њ–≤–Є–љ—Л –Є –Њ–±–ї–Њ–Љ–Ї–Є —А–∞–Ї–Њ–≤–Є–љ —В–Њ–љ–Ї–Њ—Б—В–µ–љ–љ—Л–µ, –Є–љ–Њ–≥–і–∞ - —В–µ–Љ–љ–Њ–≥–Њ —Ж–≤–µ—В–∞, –љ–µ–Њ–Ї–∞—В–∞–љ–љ—Л–µ –Є–ї–Є —Б–ї–∞–±–Њ–Њ–Ї–∞—В–∞–љ–љ—Л–µ, –њ–ї–Њ—В–љ—Л–µ, —Б—А–µ–і–љ–Є—Е –Є –љ–µ–±–Њ–ї—М—И–Є—Е —А–∞–Ј–Љ–µ—А–Њ–≤ —Б–Њ —Б–ї–µ–і–∞–Љ–Є –њ–µ—А–≤–Є—З–љ–Њ–є –Њ–Ї—А–∞—Б–Ї–Є.
–Т–Њ–Ј—А–∞—Б—В. –Ь–Є–Њ—Ж–µ–љ-–≥–Њ–ї–Њ—Ж–µ–љ. –Ш–Ј-–Ј–∞ –Њ—В–љ–Њ—Б–Є—В–µ–ї—М–љ–Њ —Б–ї–∞–±–Њ–є —Д–Њ—Б—Б–Є–ї–Є–Ј–∞—Ж–Є–Є, –љ–µ–Њ–њ–ї–µ–є—Б—В–Њ—Ж–µ–љ. –Э–∞–Є–±–Њ–ї–µ–µ –±–ї–Є–Ј–Ї–Є–є –Ї–Њ–Љ–њ–ї–µ–Ї—Б –≤—Б—В—А–µ—З–µ–љ –≤ —Б—А–µ–і–љ–µ–Љ –љ–µ–Њ–њ–ї–µ–є—Б—В–Њ—Ж–µ–љ–µ (—Н–њ–Њ—Е–∞ –њ–Њ—Е–Њ–ї–Њ–і–∞–љ–Є—П) (–Ъ—А—Л–ї–Њ–≤, –У—Г—Б–µ–≤, 2010).
–£—Б–ї–Њ–≤–Є—П –Њ–±–Є—В–∞–љ–Є—П. –Т–µ—А–Њ—П—В–љ–Њ, –њ–µ—Б—З–∞–љ—Л–µ –Є –≥–ї–Є–љ–Є—Б—В—Л–µ –Є–ї—Л —И–µ–ї—М—Д–∞ –Р—А–Ї—В–Є—З–µ—Б–Ї–Њ–≥–Њ –±–∞—Б—Б–µ–є–љ–∞. –Э–∞–ї–Є—З–Є–µ —А–∞–Ї–Њ–≤–Є–љ –Љ–Њ—А—Б–Ї–Є—Е –Љ–Њ–ї–ї—О—Б–Ї–Њ–≤ —Б –і–≤—Г–Љ—П —Б–Њ–Љ–Ї–љ—Г—В—Л–Љ–Є —Б—В–≤–Њ—А–Ї–∞–Љ–Є –Љ–Њ–ґ–µ—В —Б–≤–Є–і–µ—В–µ–ї—М—Б—В–≤–Њ–≤–∞—В—М –Њ–± –Є—Е –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ–Љ –Ј–∞—Е–Њ—А–Њ–љ–µ–љ–Є–Є –љ–∞ –Љ–µ—Б—В–µ –Њ–±–Є—В–∞–љ–Є—П –Є –Љ–Њ—А—Б–Ї–Њ–Љ –≥–µ–љ–µ–Ј–Є—Б–µ –≤–Љ–µ—Й–∞—О—Й–Є—Е –Є—Е –Њ—Б–∞–і–Ї–Њ–≤, –Є–Ј –Ї–Њ—В–Њ—А—Л—Е –±—Л–ї–∞ –Њ—В–Њ–±—А–∞–љ–∞ –і–∞–љ–љ–∞—П –њ—А–Њ–±–∞.
–Ю–±—А–∞–Ј–µ—Ж вДЦ 3298/15
- Mya truncata Linnaeus, 1758 - 2 –њ–Њ–≤—А–µ–ґ–і–µ–љ–љ—Л–µ —А–∞–Ї–Њ–≤–Є–љ—Л;
- Hiatella arctic–∞ (Linnaeus, 1758) вАУ 2 –Ј–∞–Љ–Ї–∞ –Є 1 –њ–Њ–≤—А–µ–ґ–і–µ–љ–љ–∞—П —А–∞–Ї–Њ–≤–Є–љ–∞.
–Ю–±—А–∞–Ј–µ—Ж вДЦ 3298/17
- Natica clausa (Broderip et Sowerby, 1829) вАУ 1 –њ–Њ–≤—А–µ–ґ–і–µ–љ–љ–∞—П —А–∞–Ї–Њ–≤–Є–љ–∞;
- Mya sp. (–≤–µ—А–Њ—П—В–љ–Њ –њ—А–Є–љ–∞–і–ї–µ–ґ–Є—В –Ї Mya —Бf. truncata Linnaeus, 1758) -1 –Њ–±–ї–Њ–Љ–Њ–Ї —А–∞–Ї–Њ–≤–Є–љ—Л;
- Buccinum sp. вАУ 1 –Њ–±–ї–Њ–Љ–Њ–Ї —А–∞–Ї–Њ–≤–Є–љ—Л;
- –Э–µ–Њ–њ—А–µ–і–µ–ї–Є–Љ—Л–µ –Њ–±–ї–Њ–Љ–Ї–Є —А–∞–Ї–Њ–≤–Є–љ –Љ–Њ–ї–ї—О—Б–Ї–Њ–≤ вАУ 3 —И—В.
–С—Г–Ї—Ж–Є–љ—Г–Љ—Л вАУ –±–µ–љ—В–Њ—Б–љ—Л–µ –Њ–±–Є—В–∞—В–µ–ї–Є –∞—А–Ї—В–Є—З–µ—Б–Ї–Є—Е –Љ–Њ—А–µ–є. –Ю–±–Є—В–∞—О—В –Ї–∞–Ї –љ–∞ –Љ—П–≥–Ї–Є—Е –≥–ї–Є–љ–Є—Б—В—Л—Е, –њ–µ—Б—З–∞–љ–Њ-–≥–ї–Є–љ–Є—Б—В—Л—Е –Є –њ–µ—Б—З–∞–љ–Є—Б—В—Л—Е —В–∞–Ї –Є —В–≤–µ—А–і—Л—Е –Ї–∞–Љ–µ–љ–Є—Б—В—Л—Е –≥—А—Г–љ—В–∞—Е –љ–∞ –≥–ї—Г–±–Є–љ–∞—Е 5вАУ300 –Љ (—А–µ–і–Ї–Њ –±–Њ–ї–µ–µ) –њ—А–Є —Б–Њ–ї–µ–љ–Њ—Б—В–Є 25,0вАУ35,5вА∞ –Є –љ–Є–Ј–Ї–Є—Е -2вАУ12o–° —В–µ–Љ–њ–µ—А–∞—В—Г—А–∞—Е (–Я–µ—В—А–Њ–≤, 1966, –Э–∞—Г–Љ–Њ–≤, 2006). Natica clausa (Broderip et Sowerby, 1829) - –±–Њ—А–µ–∞–ї—М–љ–Њ-–∞—А–Ї—В–Є—З–µ—Б–Ї–Є–є –≤–Є–і –Њ–±–Є—В–∞–µ—В –≤ –Р—В–ї–∞–љ—В–Є—З–µ—Б–Ї–Њ–Љ –Њ–Ї–µ–∞–љ–µ –љ–∞ —О–≥ –і–Њ –Љ—Л—Б–∞ –У–∞—В—В–µ—А–∞—Б –њ–Њ –Р–Љ–µ—А–Є–Ї–∞–љ—Б–Ї–Њ–Љ—Г –Є –Я–Њ—А—В—Г–≥–∞–ї–Є–Є –њ–Њ –Х–≤—А–Њ–њ–µ–є—Б–Ї–Њ–Љ—Г –њ–Њ–±–µ—А–µ–ґ—М—О, –≤ –Ј–∞–њ–∞–і–љ–Њ–є —З–∞—Б—В–Є –°—А–µ–і–Є–Ј–µ–Љ–љ–Њ–≥–Њ –Љ–Њ—А—П, –≤ –°–µ–≤–µ—А–љ–Њ–Љ –Ы–µ–і–Њ–≤–Є—В–Њ–Љ –Њ–Ї–µ–∞–љ–µ –љ–∞ –Ј–∞–њ–∞–і–µ –і–Њ –Ј–∞–њ–∞–і–љ–Њ–є –Є –≤–Њ—Б—В–Њ—З–љ–Њ–є –У—А–µ–љ–ї–∞–љ–і–Є–Є, –љ–∞ —Б–µ–≤–µ—А–Њ-–Ј–∞–њ–∞–і–µ –і–Њ –Ч–µ–Љ–ї–Є –§—А–∞–љ—Ж–∞-–Ш–Њ—Б–Є—Д–∞ –Є –∞—А—Е. –®–њ–Є—Ж–±–µ—А–≥–µ–љ –≤ –≤–Њ—Б—В–Њ—З–љ–Њ–є —З–∞—Б—В–Є –Љ–Њ—А—П –Ы–∞–њ—В–µ–≤—Л—Е, –≤ –Ґ–Є—Е–Њ–Љ –Њ–Ї–µ–∞–љ–µ –Њ—В –С–µ—А–Є–љ–≥–Њ–≤–∞ –њ—А–Њ–ї–Є–≤–∞ –њ–Њ –∞–Ј–Є–∞—В—Б–Ї–Њ–Љ—Г –њ–Њ–±–µ—А–µ–ґ—М—О –љ–∞ —О–≥ –і–Њ —Б–µ–≤–µ—А–љ–Њ–є —З–∞—Б—В–Є –ѓ–њ–Њ–љ—Б–Ї–Њ–≥–Њ –Љ–Њ—А—П, –њ–Њ –∞–Љ–µ—А–Є–Ї–∞–љ—Б–Ї–Њ–Љ—Г - –љ–∞ —О–≥ –і–Њ –Њ-–≤–∞ –Т–∞–љ–Ї—Г–≤–µ—А –Њ—В –љ–Є–ґ–љ–µ–є –ї–Є—В–Њ—А–∞–ї–Є (5 –Љ) –і–Њ –≥–ї—Г–±–Є–љ—Л 2660 –Љ. –њ—А–µ–Є–Љ—Г—Й–µ—Б—В–≤–µ–љ–љ–Њ –љ–∞ –Є–ї–Є—Б—В—Л—Е –Є –Є–ї–Є—Б—В–Њ-–њ–µ—Б—З–∞–љ—Л—Е –≥—А—Г–љ—В–∞—Е, –њ—А–Є —В–µ–Љ–њ–µ—А–∞—В—Г—А–µ –≤–Њ–і—Л –Њ—В -1,4 –і–Њ 16o–° –Є —Б–Њ–ї–µ–љ–Њ—Б—В–Є 25,0вАУ35,5вА∞ (–Я–µ—В—А–Њ–≤, 1966, –Э–∞—Г–Љ–Њ–≤, 2006). –Ь–Є–Є вАУ –Њ–±–Є—В–∞—В–µ–ї–Є –Љ—П–≥–Ї–Є—Е –Є–ї–Є—Б—В—Л—Е –Є–ї–Є –њ–µ—Б—З–∞–љ–Њ вАУ –Є–ї–Є—Б—В—Л—Е –Є–ї–Є —А–µ–ґ–µ –њ–µ—Б—З–∞–љ—Л—Е –≥—А—Г–љ—В–Њ–≤ –Љ–Њ—А–µ–є –°–µ–≤–µ—А–љ–Њ–≥–Њ –Ы–µ–і–Њ–≤–Є—В–Њ–≥–Њ –Њ–Ї–µ–∞–љ–∞ –Є –Љ–Њ—А–µ–є —Б–µ–≤–µ—А–∞ –Р—В–ї–∞–љ—В–Є—З–µ—Б–Ї–Њ–≥–Њ –Є –Ґ–Є—Е–Њ–≥–Њ –Њ–Ї–µ–∞–љ–Њ–≤ –≤ –Љ–Є–Њ—Ж–µ–љ–µ-–≥–Њ–ї–Њ—Ж–µ–љ–µ (https://en.wikipedia.org/wiki/Mya truncata # /media/File:Mya-truncata_in-situ_hg.jpg).
–°–Њ—Е—А–∞–љ–љ–Њ—Б—В—М. –†–∞–Ї–Њ–≤–Є–љ–∞ –Є –Њ–±–ї–Њ–Љ–Ї–Є —А–∞–Ї–Њ–≤–Є–љ —В–Њ–љ–Ї–Њ—Б—В–µ–љ–љ—Л–µ, –Њ–±–µ–ї–µ–љ—Л, –њ–ї–Њ—В–љ—Л–є, –Љ–µ–ї–Њ–≤–∞–љ–љ—Л–µ, –љ–µ–Њ–Ї–∞—В–∞–љ–љ—Л–µ, –ї–µ–≥–Ї–Є–µ, –љ–µ–±–Њ–ї—М—И–Є—Е —А–∞–Ј–Љ–µ—А–Њ–≤, —Б–ї–∞–±–Њ—Д–Њ—Б—Б–Є–ї–Є–Ј–Є—А–Њ–≤–∞–љ—Л–µ –Є–љ–Њ–≥–і–∞ —Б–Њ —Б–ї–µ–і–∞–Љ–Є –њ–µ—А–≤–Є—З–љ–Њ–є –Њ–Ї—А–∞—Б–Ї–Є.
–Т–Њ–Ј—А–∞—Б—В. –Ь–Є–Њ—Ж–µ–љ-–≥–Њ–ї–Њ—Ж–µ–љ. –Ш–Ј-–Ј–∞ –Њ—В–љ–Њ—Б–Є—В–µ–ї—М–љ–Њ —Б–ї–∞–±–Њ–є —Д–Њ—Б—Б–Є–ї–Є–Ј–∞—Ж–Є–Є, –≤–µ—А–Њ—П—В–љ–Њ, –љ–µ–Њ–њ–ї–µ–є—Б—В–Њ—Ж–µ–љ, –љ–∞–ї–Є—З–Є–µ –±—Г–Ї—Ж–Є–љ—Г–Љ–∞ –≥–Њ–≤–Њ—А–Є—В –Њ –≤–Њ–Ј–Љ–Њ–ґ–љ–Њ—Б—В–Є –њ—А–Є–љ–∞–і–ї–µ–ґ–љ–Њ—Б—В–Є —Н—В–Њ–≥–Њ –Ї–Њ–Љ–њ–ї–µ–Ї—Б–∞ –Ї —Н–њ–Њ—Е–µ –њ–Њ—В–µ–њ–ї–µ–љ–Є—П (–Ъ—А—Л–ї–Њ–≤, –Я–µ–љ–і–µ–ї—П–Ї, –Ъ–Њ—Б—В–Є–љ, 2019). –С–Њ–ї–µ–µ —В–Њ—З–љ–Њ–µ –Њ–њ—А–µ–і–µ–ї–µ–љ–Є–µ –≤–Њ–Ј—А–∞—Б—В–∞ –љ–µ –њ—А–µ–і—Б—В–∞–≤–ї—П–µ—В—Б—П –≤–Њ–Ј–Љ–Њ–ґ–љ—Л–Љ.
–£—Б–ї–Њ–≤–Є—П –Њ–±–Є—В–∞–љ–Є—П. –Т–µ—А–Њ—П—В–љ–Њ, –њ–µ—Б—З–∞–љ—Л–µ –Є –≥–ї–Є–љ–Є—Б—В—Л–µ –Є–ї—Л —И–µ–ї—М—Д–∞ –Р—А–Ї—В–Є—З–µ—Б–Ї–Њ–≥–Њ –±–∞—Б—Б–µ–є–љ–∞.
–§–Њ—В–Њ–≥—А–∞—Д–Є–Є –Ї–Њ–ї–ї–µ–Ї—Ж–Є–Є –Є–Ј—Г—З–µ–љ–љ—Л—Е –Љ–Њ—А—Б–Ї–Є—Е –Љ–Њ–ї–ї—О—Б–Ї–Њ–≤ –°–µ–≤–µ—А–љ–Њ–є –Ч–µ–Љ–ї–Є (–Њ. –С–Њ–ї—М—И–µ–≤–Є–Ї), —Б–±–Њ—А—Л 2020 –≥.


–§–Њ—В–Њ 1вАУ2. –І–µ—В—Л—А–µ —А–∞–Ї–Њ–≤–Є–љ—Л –Љ–Њ—А—Б–Ї–Є—Е –і–≤—Г—Б—В–≤–Њ—А—З–∞—В—Л—Е –Љ–Њ–ї–ї—О—Б–Ї–Њ–≤ Mya truncata Linnaeus, 1758 –Є–Ј –њ—А–Њ–±—Л 3298/7 (—Б –і–≤—Г–Љ—П —Б–Њ–Љ–Ї–љ—Г—В—Л–Љ–Є —Б—В–≤–Њ—А–Ї–∞–Љ–Є –Є –Ї–Њ–љ—Е–Є–Њ–ї–Є–љ–љ–Њ–≤—Л–Љ —Б–ї–Њ–µ–Љ), –Ї–Њ—В–Њ—А—Л–µ —Б–≤–Є–і–µ—В–µ–ї—М—Б—В–≤—Г—О—В –Њ –њ—А–Є–ґ–Є–Ј–љ–µ–љ–љ–Њ–Љ –Ј–∞—Е–Њ—А–Њ–љ–µ–љ–Є–Є (–≤–Є–і —Б–≤–µ—А—Е—Г –Є —Б–љ–Є–Ј—Г). –Ф–ї–Є–љ–∞ –љ–Є–ґ–љ–µ–є –њ—А–∞–≤–Њ–є —А–∞–Ї–Њ–≤–Є–љ—Л 2,5 —Б–Љ

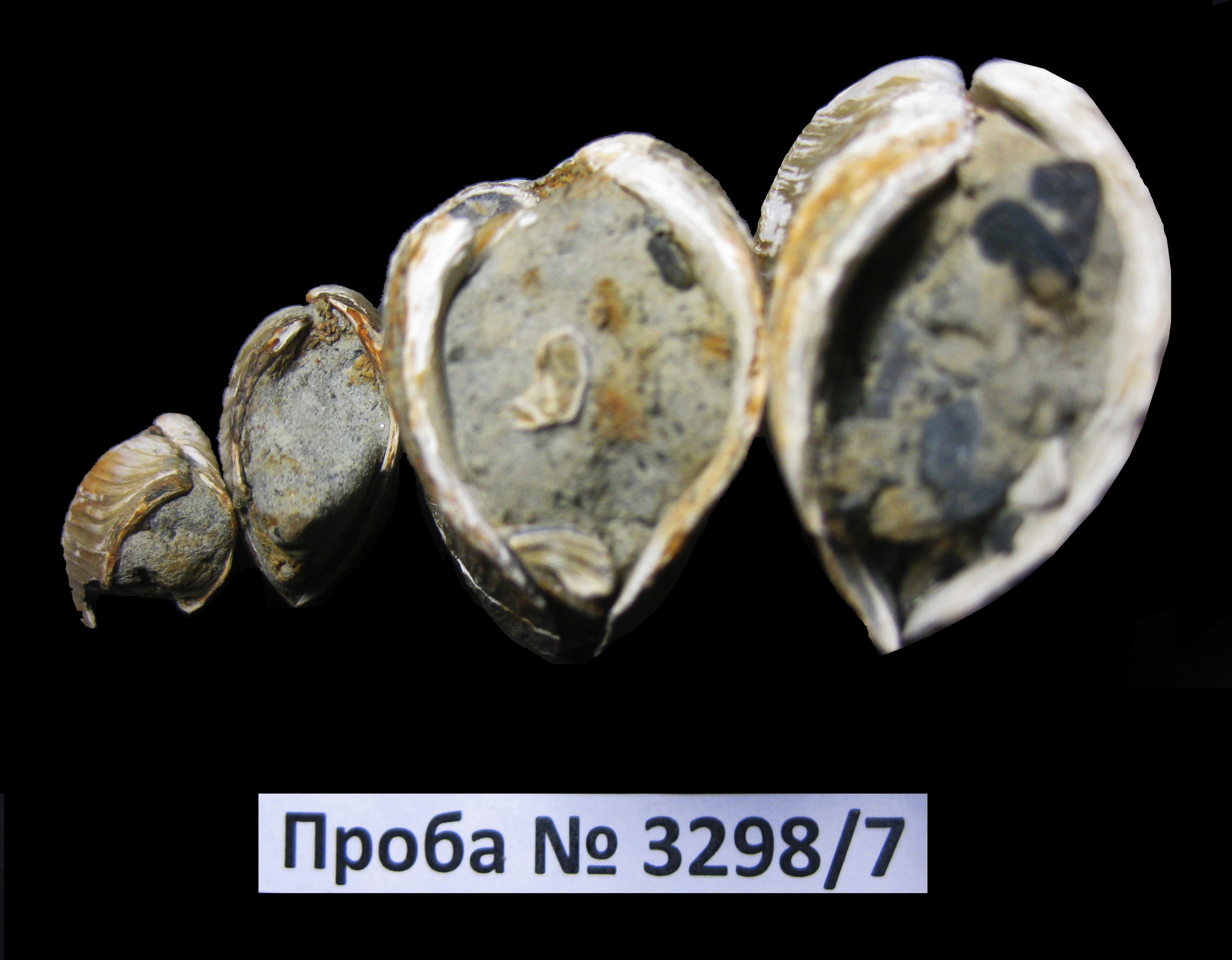
–§–Њ—В–Њ 3вАУ4. –І–µ—В—Л—А–µ —А–∞–Ї–Њ–≤–Є–љ—Л –Љ–Њ—А—Б–Ї–Є—Е –і–≤—Г—Б—В–≤–Њ—А—З–∞—В—Л—Е –Љ–Њ–ї–ї—О—Б–Ї–Њ–≤ Mya truncata Linnaeus, 1758 –Є–Ј –њ—А–Њ–±—Л 3298/7 (—Б –і–≤—Г–Љ—П —Б–Њ–Љ–Ї–љ—Г—В—Л–Љ–Є —Б—В–≤–Њ—А–Ї–∞–Љ–Є –Є –Ї–Њ–љ—Е–Є–Њ–ї–Є–љ–љ–Њ–≤—Л–Љ —Б–ї–Њ–µ–Љ), –Ї–Њ—В–Њ—А—Л–µ —Б–≤–Є–і–µ—В–µ–ї—М—Б—В–≤—Г—О—В –Њ –њ—А–Є–ґ–Є–Ј–љ–µ–љ–љ–Њ–Љ –Ј–∞—Е–Њ—А–Њ–љ–µ–љ–Є–Є (–≤–Є–і —Б–ї–µ–≤–∞ –Є —Б–њ—А–∞–≤–∞). –®–Є—А–Є–љ–∞ –ї–µ–≤–Њ–є —А–∞–Ї–Њ–≤–Є–љ—Л 1,2 —Б–Љ

–§–Њ—В–Њ 5. –Ь–љ–Њ–≥–Њ—З–Є—Б–ї–µ–љ–љ—Л–µ –њ–Њ–≤—А–µ–ґ–і–µ–љ–љ—Л–µ —А–∞–Ї–Њ–≤–Є–љ—Л –Љ–Њ—А—Б–Ї–Є—Е –і–≤—Г—Б—В–≤–Њ—А—З–∞—В—Л—Е –Љ–Њ–ї–ї—О—Б–Ї–Њ–≤ Mya truncata Linnaeus, 1758 –Є –Є—Е –Њ–±–ї–Њ–Љ–Ї–Є –Є–Ј –њ—А–Њ–±—Л 3298/7 –Є –Ї–Њ–љ—Е–Є–Њ–ї–Є–љ–љ–Њ–≤—Л–Љ —Б–ї–Њ–µ–Љ (–≤–Є–і —Б–≤–µ—А—Е—Г). –Ф–ї–Є–љ–∞ –њ–Њ–≤—А–µ–ґ–і–µ–љ–љ–Њ–є –љ–Є–ґ–љ–µ–є –ї–µ–≤–Њ–є —А–∞–Ї–Њ–≤–Є–љ—Л 1,5 —Б–Љ

–§–Њ—В–Њ 6. –Ь–љ–Њ–≥–Њ—З–Є—Б–ї–µ–љ–љ—Л–µ –њ–Њ–≤—А–µ–ґ–і–µ–љ–љ—Л–µ —А–∞–Ї–Њ–≤–Є–љ—Л –Љ–Њ—А—Б–Ї–Є—Е –і–≤—Г—Б—В–≤–Њ—А—З–∞—В—Л—Е –Љ–Њ–ї–ї—О—Б–Ї–Њ–≤ Mya truncata Linnaeus, 1758 –Є –Є—Е –Њ–±–ї–Њ–Љ–Ї–Є –Є–Ј –њ—А–Њ–±—Л 3298/7 (–≤–Є–і —Б–љ–Є–Ј—Г). –Ф–ї–Є–љ–∞ –њ–Њ–≤—А–µ–ґ–і–µ–љ–љ–Њ–є –љ–Є–ґ–љ–µ–є –ї–µ–≤–Њ–є —А–∞–Ї–Њ–≤–Є–љ—Л 1,5 —Б–Љ


–§–Њ—В–Њ 7вАУ8. –†–∞–Ї–Њ–≤–Є–љ—Л –Љ–Њ—А—Б–Ї–Є—Е –Љ–Њ–ї–ї—О—Б–Ї–Њ–≤ Mya truncata Linnaeus, 1758 –Є Hiatella arctica (Linaneus, 1758) –Є–Ј –њ—А–Њ–±—Л вДЦ 3298/15. –Ф–ї–Є–љ–∞ –ї–µ–≤–Њ–є —А–∞–Ї–Њ–≤–Є–љ—Л 3 —Б–Љ


–§–Њ—В–Њ 9вАУ10. –†–∞–Ї–Њ–≤–Є–љ–∞ –Љ–Њ—А—Б–Ї–Њ–≥–Њ –Љ–Њ–ї–ї—О—Б–Ї–∞ Natica clausa (Broderip et Sowerby, 1824) —Б–Њ —Б–ї–µ–і–∞–Љ–Є –њ–µ—А–≤–Є—З–љ–Њ–є –Њ–Ї—А–∞—Б–Ї–Є –Є –Њ–±–ї–Њ–Љ–Ї–Є —А–∞–Ї–Њ–≤–Є–љ –Љ–Њ—А—Б–Ї–Є—Е –Љ–Њ–ї–ї—О—Б–Ї–Њ–≤ Natica clausa (Broderip et Sowerby, 1829), Mya sp., Buccinum sp. –Є–Ј –њ—А–Њ–±—Л вДЦ 3298/17. –Ф–Є–∞–Љ–µ—В—А –ї–µ–≤–Њ–є —А–∞–Ї–Њ–≤–Є–љ—Л 1 —Б–Љ

–§–Њ—В–Њ 11. –†–∞–Ї–Њ–≤–Є–љ–∞ –Є –Њ–±–ї–Њ–Љ–Њ–Ї —А–∞–Ї–Њ–≤–Є–љ—Л –Љ–Њ—А—Б–Ї–Њ–≥–Њ –Љ–Њ–ї–ї—О—Б–Ї–∞ Natica clausa (Broderip et Sowerby, 1829) —Б–Њ —Б–ї–µ–і–∞–Љ–Є –њ–µ—А–≤–Є—З–љ–Њ–є –Њ–Ї—А–∞—Б–Ї–Є

–§–Њ—В–Њ 12. –Т–љ–µ—И–љ–Є–є –≤–Є–і –Њ–±–љ–∞–ґ–µ–љ–Є—П 1, –Є–Ј –Ї–Њ—В–Њ—А–Њ–≥–Њ –њ—А–Њ–Є—Б—Е–Њ–і—П—В –Є–Ј—Г—З–µ–љ–љ—Л–µ —А–∞–Ї–Њ–≤–Є–љ—Л –Љ–Њ–ї–ї—О—Б–Ї–Њ–≤. –Ю—Б—В—А–Њ–≤ –С–Њ–ї—М—И–µ–≤–Є–Ї, –∞—А—Е–Є–њ–µ–ї–∞–≥ –°–µ–≤–µ—А–љ–∞—П –Ч–µ–Љ–ї—П. –§–Њ—В–Њ –Ф. –Ѓ. –С–Њ–ї—М—И–Є—П–љ–Њ–≤–∞, 2020 –≥


–§–Њ—В–Њ 13вАУ14. –Т–љ–µ—И–љ–Є–є –≤–Є–і –Њ–±–љ–∞–ґ–µ–љ–Є—П 2 —Б –Ј–∞—Е–Њ—А–Њ–љ–µ–љ–љ—Л–Љ–Є —А–∞–Ї–Њ–≤–Є–љ–∞–Љ–Є –Љ–Є–є —Б –і–≤—Г–Љ—П —Б–Њ–Љ–Ї–љ—Г—В—Л–Љ–Є —Б—В–≤–Њ—А–Ї–∞–Љ–Є (–≤ –њ—А–Є–ґ–Є–Ј–љ–µ–љ–љ–Њ–Љ –њ–Њ–ї–Њ–ґ–µ–љ–Є–Є) –Є —Б–Є—Д–Њ–љ–∞–Љ–Є –≤–≤–µ—А—Е, —З—В–Њ —Б–≤–Є–і–µ—В–µ–ї—М—Б—В–≤—Г–µ—В –Њ –Љ–Њ—А—Б–Ї–Њ–Љ –њ—А–Њ–Є—Б—Е–Њ–ґ–і–µ–љ–Є–Є –≤–Љ–µ—Й–∞—О—Й–Є—Е –Є—Е –њ–Њ—А–Њ–і. –Ю—Б—В—А–Њ–≤ –С–Њ–ї—М—И–µ–≤–Є–Ї, –∞—А—Е–Є–њ–µ–ї–∞–≥ –°–µ–≤–µ—А–љ–∞—П –Ч–µ–Љ–ї—П. –§–Њ—В–Њ –Ф. –Ѓ. –С–Њ–ї—М—И–Є—П–љ–Њ–≤–∞, 2020 –≥
–Ю–њ—А–µ–і–µ–ї–µ–љ–Є–µ –Ї–Њ—Б—В–љ—Л—Е –Њ—Б—В–∞—В–Ї–Њ–≤ –Љ–Њ—А—Б–Ї–Є—Е –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е –њ–Њ —Д–Њ—В–Њ–≥—А–∞—Д–Є—П–Љ (—А–Є—Б. 1) –њ—А–Њ–≤—С–ї –њ–∞–ї–µ–Њ–љ—В–Њ–ї–Њ–≥ –Р. –Т. –Ъ—А—Л–ї–Њ–≤. –°—А–µ–і–Є —Б–±–Њ—А–Њ–≤ –Ї–Њ—Б—В–µ–є –Њ–Ї–∞–Ј–∞–ї–Є—Б—М –њ–Њ–Ј–≤–Њ–љ–Ї–Є —Б–µ—А–Њ–≥–Њ –Ї–Є—В–∞ Eschrichtius robustus (Lilljeborg, 1861) —А—С–±—А–∞ –Є —З–∞—Б—В—М —З–µ–ї—О—Б—В–Є –ї–∞–њ—В–µ–≤—Б–Ї–Њ–≥–Њ –Љ–Њ—А–ґ–∞ Odobenus rosmarus laptevi (Chapskii, 1940).

–†–Є—Б. 1 –°–±–Њ—А—Л –Ї–Њ—Б—В–µ–є –Љ–Њ—А—Б–Ї–Є—Е –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е, –≤—Л–Љ—Л—В—Л—Е –Є–Ј –Њ—В–ї–Њ–ґ–µ–љ–Є–є –Њ–±–љ–∞–ґ–µ–љ–Є—П 3298
–Ю—В–ї–Њ–ґ–µ–љ–Є—П –љ–∞—Е–Њ–і—П—В—Б—П –≤ —Б—В–∞–і–Є–Є –і–∞—В–Є—А–Њ–≤–∞–љ–Є—П –Є —Г–ґ–µ –њ–Њ–ї—Г—З–µ–љ—Л 2 –і–∞—В–Є—А–Њ–≤–Ї–Є. –Т –љ–∞—Г—З–љ–Њ-–Є—Б—Б–ї–µ–і–Њ–≤–∞—В–µ–ї—М—Б–Ї–Њ–є –ї–∞–±–Њ—А–∞—В–Њ—А–Є–Є –≥–µ–Њ—Е—А–Њ–љ–Њ–ї–Њ–≥–Є–Є —З–µ—В–≤–µ—А—В–Є—З–љ–Њ–≥–Њ –њ–µ—А–Є–Њ–і–∞ ¬ЂGeoAge¬ї (–Ґ–∞–ї–ї–Є–љ–љ, –≠—Б—В–Њ–љ–Є—П) –Љ–µ—В–Њ–і–Њ–Љ –≠–Я–† (—Н–ї–µ–Ї—В—А–Њ–љ–љ–Њ-–њ–∞—А–∞–Љ–∞–≥–љ–Є—В–љ–Њ-—А–µ–Ј–Њ–љ–∞–љ—Б–љ–Њ–є —Б–њ–µ–Ї—В—А–Њ—Б–Ї–Њ–њ–Є–Є) –і–∞—В–Є—А–Њ–≤–∞–љ—Л —А–∞–Ї–Њ–≤–Є–љ—Л –Љ–Њ—А—Б–Ї–Є—Е –Љ–Њ–ї–ї—О—Б–Ї–Њ–≤ –Є –≤–Љ–µ—Й–∞—О—Й–Є–µ –Є—Е —З—С—А–љ—Л–µ –≥–ї–Є–љ—Л (–Њ–±—А–∞–Ј–µ—Ж 3298/6). –Ш—Е –≤–Њ–Ј—А–∞—Б—В –Њ–Ї–∞–Ј–∞–ї—Б—П —А–∞–≤–љ—Л–Љ 95,6¬±7,4 —В—Л—Б. –ї–µ—В.
–†–∞–і–Є–Њ—Г–≥–ї–µ—А–Њ–і–љ–Њ–µ –і–∞—В–Є—А–Њ–≤–∞–љ–Є–µ –≤—Л–њ–Њ–ї–љ–µ–љ–Њ –≤ –Ы–∞–±–Њ—А–∞—В–Њ—А–Є–Є –≥–µ–Њ–Љ–Њ—А—Д–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є—Е –Є –њ–∞–ї–µ–Њ–≥–µ–Њ–≥—А–∞—Д–Є—З–µ—Б–Ї–Є—Е –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–є –њ–Њ–ї—П—А–љ—Л—Е —А–µ–≥–Є–Њ–љ–Њ–≤ –Є –Ь–Є—А–Њ–≤–Њ–≥–Њ –Њ–Ї–µ–∞–љ–∞ –Є–Љ. –Ъ—С–њ–њ–µ–љ–∞ –°–Я–±–У–£. –Ф–∞—В–Є—А–Њ–≤–∞–љ —Б—В–≤–Њ–ї –і—А–µ–≤–µ—Б–Є–љ—Л –њ–ї–∞–≤–љ–Є–Ї–∞ –Є–Ј –њ–∞—З–Ї–Є –њ–µ—Б—З–∞–љ–Њ-–≥—А–∞–≤–Є–є–љ—Л—Е –Љ–Њ—А—Б–Ї–Є—Е –Њ—Б–∞–і–Ї–Њ–≤, –њ–µ—А–µ–Ї—А—Л–≤–∞—О—Й–Є—Е –≥–ї–Є–љ—Л. –†–∞–і–Є–Њ—Г–≥–ї–µ—А–Њ–і–љ—Л–є –≤–Њ–Ј—А–∞—Б—В –Њ–±—А–∞–Ј—Ж–∞ —Б–Њ—Б—В–∞–≤–Є–ї 45760¬±1160 –ї–µ—В (–Ї–∞–ї–µ–љ–і–∞—А–љ—Л–є –≤–Њ–Ј—А–∞—Б—В 48630¬±1640 –ї–µ—В).
–Ш—Б—Б–ї–µ–і–Њ–≤–∞–љ–љ–Њ–µ –Њ–±–љ–∞–ґ–µ–љ–Є–µ –Љ–Њ—А—Б–Ї–Є—Е –Њ—Б–∞–і–Ї–Њ–≤ –љ–∞ –±–µ—А–µ–≥—Г –Ј–∞–ї–Є–≤–∞ –Р—Е–Љ–∞—В–Њ–≤–∞ –њ—А–Є–≤–љ–µ—Б–ї–Њ –љ–Њ–≤—Л–µ –і–∞–љ–љ—Л–µ –і–ї—П –њ–Њ–љ–Є–Љ–∞–љ–Є—П –Љ–Њ—А—Б–Ї–Њ–є –Є—Б—В–Њ—А–Є–Є –∞—А—Е–Є–њ–µ–ї–∞–≥–∞ –°–µ–≤–µ—А–љ–∞—П –Ч–µ–Љ–ї—П –≤ –≤–µ—А—Е–љ–µ–Љ –љ–µ–Њ–њ–ї–µ–є—Б—В–Њ—Ж–µ–љ–µ. –Ю–±–љ–∞—А—Г–ґ–µ–љ–љ—Л–µ –Є –Њ—Е–∞—А–∞–Ї—В–µ—А–Є–Ј–Њ–≤–∞–љ–љ—Л–µ —А–µ–Ј—Г–ї—М—В–∞—В–∞–Љ–Є –∞–љ–∞–ї–Є–Ј–Њ–≤ –Њ—Б–∞–і–Ї–Є —Б–≤–Є–і–µ—В–µ–ї—М—Б—В–≤—Г—О—В –Њ —В–Њ–Љ, —З—В–Њ –Љ–Њ—А—Б–Ї–Є–µ —Г—Б–ї–Њ–≤–Є—П –≥–Њ—Б–њ–Њ–і—Б—В–≤–Њ–≤–∞–ї–Є –Ј–і–µ—Б—М 95 –Є 47 —В—Л—Б—П—З –ї–µ—В –љ–∞–Ј–∞–і. –Я—А–Є—З—С–Љ –Љ–Њ—А–µ –≤ –Њ–±–∞ –њ–µ—А–Є–Њ–і–∞ –±—Л–ї–Њ –Њ—В–Ї—А—Л—В—Л–Љ –Њ—В –ї–µ–і—П–љ–Њ–≥–Њ –њ–Њ–Ї—А–Њ–≤–∞, —В.–µ. –Є–Љ–µ–ї–Њ —Б–µ–Ј–Њ–љ–љ—Л–є –њ–Њ–Ї—А–Њ–≤ –ї—М–і–∞ –≤ —Е–Њ–ї–Њ–і–љ–Њ–µ –≤—А–µ–Љ—П –≥–Њ–і–∞, –Ї–∞–Ї –Є –≤ –љ–∞—Б—В–Њ—П—Й–µ–µ –≤—А–µ–Љ—П. –Ґ–µ–Љ–њ–µ—А–∞—В—Г—А—Л –њ—А–Є–і–Њ–љ–љ–Њ–є –≤–Њ–і—Л –±—Л–ї–Є –Њ—В—А–Є—Ж–∞—В–µ–ї—М–љ—Л–µ –Є–ї–Є –±–ї–Є–Ј–Ї–Є–Љ–Є –Ї 0¬∞–°, —Б–Њ–ї—С–љ–Њ—Б—В—М –≤–Њ–і—Л –±–ї–Є–Ј–Ї–Њ–є –Ї –љ–Њ—А–Љ–∞–ї—М–љ–Њ–є. –≠—В–Њ –њ–µ—А–Є–Њ–і –љ–µ –±—Л–ї –≤—А–µ–Љ–µ–љ–µ–Љ —А–∞–Ј–≤–Є—В–Є—П –њ–Њ–Ї—А–Њ–≤–љ–Њ–≥–Њ –Њ–ї–µ–і–µ–љ–µ–љ–Є—П. –С–Њ–ї–µ–µ —В–Њ–≥–Њ, –і–∞–ґ–µ —В–µ –ї–µ–і–љ–Є–Ї–Є, –Ї–Њ—В–Њ—А—Л–µ —А–∞–Ј–≤–Є–≤–∞–ї–Є—Б—М –љ–∞ –∞—А—Е–Є–њ–µ–ї–∞–≥–µ, —З–∞—Б—В–Њ –љ–µ –і–Њ—Б—В–Є–≥–∞–ї–Є –±–µ—А–µ–≥–Њ–≤–Њ–є –ї–Є–љ–Є–Є –Љ–Њ—А—П. –Т –Њ–њ–Є—Б–∞–љ–љ–Њ–Љ –Њ–±–љ–∞–ґ–µ–љ–Є–Є –љ–µ—В –њ—А–Є–Ј–љ–∞–Ї–Њ–≤ –ї–µ–і–љ–Є–Ї–Њ–≤—Л—Е –Њ—В–ї–Њ–ґ–µ–љ–Є–є. –Ъ–∞–Ї 46, —В–∞–Ї –Є 95 —В—Л—Б—П—З –ї–µ—В –љ–∞–Ј–∞–і –Ј–і–µ—Б—М –≥–Њ—Б–њ–Њ–і—Б—В–≤–Њ–≤–∞–ї–Є –Њ–±—Б—В–∞–љ–Њ–≤–Ї–Є –њ—А–Є–±—А–µ–ґ–љ–Њ-–Љ–Њ—А—Б–Ї–Њ–≥–Њ –Њ—Б–∞–і–Ї–Њ–љ–∞–Ї–Њ–њ–ї–µ–љ–Є—П, —З—В–Њ –њ–Њ–і—В–≤–µ—А–ґ–і–∞–µ—В –≤—Л–≤–Њ–і—Л, –њ–Њ–ї—Г—З–µ–љ–љ—Л–µ –њ–Њ —А–µ–Ј—Г–ї—М—В–∞—В–∞–Љ –Є–Ј—Г—З–µ–љ–Є—П –Љ–Њ—А—Б–Ї–Є—Е –Њ—В–ї–Њ–ґ–µ–љ–Є–є –і—А—Г–≥–Є—Е –Њ—Б—В—А–Њ–≤–Њ–≤ –∞—А—Е–Є–њ–µ–ї–∞–≥–∞ [–С–Њ–ї—М—И–Є—П–љ–Њ–≤, –Ь–∞–Ї–µ–µ–≤, 1995] –Є –љ–Є–Ї–∞–Ї –љ–µ –њ–Њ–і—В–≤–µ—А–ґ–і–∞–µ—В –≤—Л–≤–Њ–і–Њ–≤ —И–≤–µ–і—Б–Ї–Є—Е –Є—Б—Б–ї–µ–і–Њ–≤–∞—В–µ–ї–µ–є –Њ —А–∞—Б–њ—А–Њ—Б—В—А–∞–љ–µ–љ–Є–Є –≤ –љ–∞—З–∞–ї–µ –њ–Њ–Ј–і–љ–µ–≥–Њ –љ–µ–Њ–њ–ї–µ–є—Б—В–Њ—Ж–µ–љ–∞ (90-100 —В—Л—Б. –ї–µ—В –љ–∞–Ј–∞–і –Љ–∞–Ї—Б–Є–Љ–∞–ї—М–љ–Њ–µ —А–∞—Б–њ—А–Њ—Б—В—А–∞–љ–µ–љ–Є–µ) –ї–µ–і–љ–Є–Ї–Њ–≤—Л—Е —Й–Є—В–Њ–≤ –љ–∞ –≤—Б–µ–є —В–µ—А—А–Є—В–Њ—А–Є–µ–є –∞—А—Е–Є–њ–µ–ї–∞–≥–∞ –°–µ–≤–µ—А–љ–∞—П –Ч–µ–Љ–ї—П –Є –≤ —Б–µ–≤–µ—А–љ–Њ–є —З–∞—Б—В–Є –њ-–Њ–≤–∞ –Ґ–∞–є–Љ—Л—А [Moller et al., 2015]. –Ъ—А–Њ–Љ–µ —В–Њ–≥–Њ, –≤–Њ –≤—В–Њ—А–Њ–є –њ–Њ–ї–Њ–≤–Є–љ–µ –њ–Њ–Ј–і–љ–µ–≥–Њ –љ–µ–Њ–њ–ї–µ–є—Б—В–Њ—Ж–µ–љ–∞, –∞ –Є–Љ–µ–љ–љ–Њ 46 —В—Л—Б—П—З –ї–µ—В –љ–∞–Ј–∞–і (–Ї–∞—А–≥–Є–љ—Б–Ї–∞—П —В—А–∞–љ—Б–≥—А–µ—Б—Б–Є—П –≤ –њ–Њ–љ–Є–Љ–∞–љ–Є–Є –Т.–Э. –°–∞–Ї—Б–∞ [1947]), –Ї–∞–Ї –њ–Њ–Ї–∞–Ј—Л–≤–∞–µ—В —А–∞–і–Є–Њ—Г–≥–ї–µ—А–Њ–і–љ–∞—П –і–∞—В–Є—А–Њ–≤–Ї–∞ –њ–ї–∞–≤–љ–Є–Ї–Њ–≤–Њ–є –і—А–µ–≤–µ—Б–Є–љ—Л –Є–Ј –Є–Ј—Г—З–∞–µ–Љ–Њ–≥–Њ –Њ–±–љ–∞–ґ–µ–љ–Є—П, —Г—А–Њ–≤–µ–љ—М –Љ–Њ—А—П –±—Л–ї –љ–∞ 40-60 –Љ –≤—Л—И–µ —Б–Њ–≤—А–µ–Љ–µ–љ–љ–Њ–≥–Њ. –Ш —Н—В–Њ –Љ–Њ—А–µ —В–∞–Ї–ґ–µ –±—Л–ї–Њ —Б–µ–Ј–Њ–љ–љ–Њ –Њ—В–Ї—А—Л—В—Л–Љ –Њ—В –ї—М–і–∞ —Б –Њ–±–Є—В–∞–љ–Є–µ–Љ –≤ –љ—С–Љ –Љ–Њ—А—Б–Ї–Є—Е –Љ–ї–µ–Ї–Њ–њ–Є—В–∞—О—Й–Є—Е (–Ї–Є—В–Њ–≤, –Љ–Њ—А–ґ–µ–є, –љ–∞—А–≤–∞–ї–Њ–≤), –Ї–Њ—Б—В–Є –Ї–Њ—В–Њ—А—Л—Е –і–∞—В–Є—А—Г—О—В—Б—П —А–∞–і–Є–Њ—Г–≥–ї–µ—А–Њ–і–љ—Л–Љ –≤–Њ–Ј—А–∞—Б—В–Њ–Љ 41вАУ43 —В—Л—Б—П—З–Є –ї–µ—В –љ–∞ –і—А—Г–≥–Є—Е –Њ—Б—В—А–Њ–≤–∞—Е –∞—А—Е–Є–њ–µ–ї–∞–≥–∞ [–С–Њ–ї—М—И–Є—П–љ–Њ–≤, –Ь–∞–Ї–µ–µ–≤, 1995].
- –С–Њ–ї—М—И–Є—П–љ–Њ–≤ –Ф.–Ѓ., –Ь–∞–Ї–µ–µ–≤ –Т. –Э. –Р—А—Е–Є–њ–µ–ї–∞–≥ –°–µ–≤–µ—А–љ–∞—П –Ч–µ–Љ–ї—П. –Ю–ї–µ–і–µ–љ–µ–љ–Є–µ, –Є—Б—В–Њ—А–Є—П —А–∞–Ј–≤–Є—В–Є—П –њ—А–Є—А–Њ–і–љ–Њ–є —Б—А–µ–і—Л. –°–Я–±. –У–Є–і—А–Њ–Љ–µ—В–µ–Њ–Є–Ј–і–∞—В, 1995, 207 —Б.
- –Т–≤–µ–і–µ–љ–Є–µ –≤ –Є–Ј—Г—З–µ–љ–Є–µ —Д–Њ—А–∞–Љ–Є–љ–Є—Д–µ—А. –Я–Њ–і —А–µ–і–∞–Ї—Ж–Є–µ–є –Э. –Э. –°—Г–±–±–Њ—В–Є–љ–Њ–є, –Э. –Р. –Т–Њ–ї–Њ—И–Є–љ–Њ–≤–Њ–є, –Р.–ѓ. –Р–Ј–±–µ–ї—М, 1991–≥.
- –Ч–µ–љ–Ї–µ–≤–Є—З –Ы. –Р. –Ш–Ј–±—А–∞–љ–љ—Л–µ —В—А—Г–і—Л. –Ґ. 1: –С–Є–Њ–ї–Њ–≥–Є—П —Б–µ–≤–µ—А–љ—Л—Е –Є —О–ґ–љ—Л—Е –Љ–Њ—А–µ–є –°–°–°–†. –Ь.: –Э–∞—Г–Ї–∞, 1977, 339 —Б.
- –Ъ—А—Л–ї–Њ–≤ –Р. –Т., –У—Г—Б–µ–≤ –Х. –Р. –Ъ–Њ–Љ–њ–ї–µ–Ї—Б—Л –њ–Њ–Ј–і–љ–µ–Ї–∞–є–љ–Њ–Ј–Њ–є—Б–Ї–Є—Е –Љ–Њ–ї–ї—О—Б–Ї–Њ–≤ –Є–Ј —В–µ—А—А–∞—Б –°–µ–≤–µ—А–љ–Њ–є –Ч–µ–Љ–ї–Є // –У–µ–Њ–ї–Њ–≥–Њ-–≥–µ–Њ—Д–Є–Ј–Є—З–µ—Б–Ї–Є–µ —Е–∞—А–∞–Ї—В–µ—А–Є—Б—В–Є–Ї–Є –ї–Є—В–Њ—Б—Д–µ—А—Л –Р—А–Ї—В–Є—З–µ—Б–Ї–Њ–≥–Њ —А–µ–≥–Є–Њ–љ–∞. 2010. –Т—Л–њ. 7. –Ґ—А. –Т–Э–Ш–Ш–Ю–Ї–µ–∞–љ–≥–µ–Њ–ї–Њ–≥–Є—П. –Ґ–Њ–Љ 210, —Б. 82вАУ95.
- –Ъ—А—Л–ї–Њ–≤ –Р. –Т., –Я–µ–љ–і–µ–ї—П–Ї –†.–Э., –Ъ–Њ—Б—В–Є–љ –Ф.–Э. –Э–Њ–≤—Л–µ –і–∞–љ–љ—Л–µ –њ–Њ —Б—В—А–∞—В–Є–≥—А–∞—Д–Є—З–µ—Б–Ї–Њ–Љ—Г –Ј–љ–∞—З–µ–љ–Є—О –Ї–Њ–Љ–њ–ї–µ–Ї—Б–Њ–≤ –Љ–Њ—А—Б–Ї–Є—Е –Љ–Њ–ї–ї—О—Б–Ї–Њ–≤ –Є —А–∞–Ї–Њ–Њ–±—А–∞–Ј–љ—Л—Е –≤—В–Њ—А–Њ–є –Є —З–µ—В–≤–µ—А—В–Њ–є –Љ–Њ—А—Б–Ї–Є—Е —В–µ—А—А–∞—Б (–Ь–Ш–°-3 –Є –Ь–Ш–°-5) –њ–Њ–Ј–і–љ–µ–≥–Њ –љ–µ–Њ–њ–ї–µ–є—Б—В–Њ—Ж–µ–љ–∞ (–Ї–∞—А–≥–Є–љ—Б–Ї–Њ–є –Є –Ї–∞–Ј–∞–љ—Ж–µ–≤—Б–Ї–Њ–є —В—А–∞–љ—Б–≥—А–µ—Б—Б–Є–є) –њ-–Њ–≤–∞ –Ґ–∞–є–Љ—Л—А, –Я–∞–є-–•–Њ—П –Є –С–µ–ї–Њ–Љ–Њ—А—Б–Ї–Њ–≥–Њ-–Ъ—Г–ї–Њ–є—Б–Ї–Њ–≥–Њ –њ–ї–∞—В–Њ // –†–µ–ї—М–µ—Д –Є —З–µ—В–≤–µ—А—В–Є—З–љ—Л–µ –Њ–±—А–∞–Ј–Њ–≤–∞–љ–Є—П –Р—А–Ї—В–Є–Ї–Є, –°—Г–±–∞—А–Ї—В–Є–Ї–Є –Є –°–µ–≤–µ—А–Њ-–Ч–∞–њ–∞–і–∞ –†–Њ—Б—Б–Є–Є. –Ь–∞—В–µ—А–Є–∞–ї—Л –µ–ґ–µ–≥–Њ–і–љ–Њ–є –Ї–Њ–љ—Д–µ—А–µ–љ—Ж–Є–Є –њ–Њ —А–µ–Ј—Г–ї—М—В–∞—В–∞–Љ —Н–Ї—Б–њ–µ–і–Є—Ж–Є–Њ–љ–љ—Л—Е –Є—Б—Б–ї–µ–і–Њ–≤–∞–љ–Є–є. –Т—Л–њ. 5, –°–Я–±., 2019, —Б. 81вАУ91.
- –Э–∞—Г–Љ–Њ–≤ –Р. –Ф. –Ф–≤—Г—Б—В–≤–Њ—А—З–∞—В—Л–µ –Љ–Њ–ї–ї—О—Б–Ї–Є –±–µ–ї–Њ–≥–Њ –Љ–Њ—А—П. –≠–Ї–Њ–ї–Њ–≥–Њ-—Д–∞—Г–љ–Є—Б—В–Є—З–µ—Б–Ї–Є–є –∞–љ–∞–ї–Є–Ј. –°–Я–±.: –Ч–Њ–Њ–ї–Њ–≥–Є—З–µ—Б–Ї–Є–є –Є–љ—Б—В–Є—В—Г—В –†–Р–Э, 2006, 367 —Б.
- –Я–µ—В—А–Њ–≤ –Ю. –Ь. –°—В—А–∞—В–Є–≥—А–∞—Д–Є—П –Є —Д–∞—Г–љ–∞ –Љ–Њ—А—Б–Ї–Є—Е –Љ–Њ–ї–ї—О—Б–Ї–Њ–≤ —З–µ—В–≤–µ—А—В–Є—З–љ—Л—Е –Њ—В–ї–Њ–ґ–µ–љ–Є–є –І—Г–Ї–Њ—В—Б–Ї–Њ–≥–Њ –њ–Њ–ї—Г–Њ—Б—В—А–Њ–≤–∞. –Ґ—А—Г–і—Л –У–Ш–Э –Р–Э –°–°–°–†, –≤—Л–њ. 155. –Ь–Њ—Б–Ї–≤–∞. ¬Ђ–Э–∞—Г–Ї–∞¬ї, 1966, 252 —Б.
- –°–∞–Ї—Б –Т.–Э. –Ъ–Њ–ї–µ–±–∞–љ–Є—П —Г—А–Њ–≤–љ—П –Љ–Њ—А—П –≤ —Г—Б—В—М–µ –Х–љ–Є—Б–µ—П –≤ —З–µ—В–≤–µ—А—В–Є—З–љ—Л–є –њ–µ—А–Є–Њ–і // –Я—А–Є—А–Њ–і–∞. 1947. вДЦ 5. –°. 51вАУ54.
- McNeil F. S. Evolution and distribution of the genus Mya and Tertiary migrations of Mollusca Professional Paper. 1965, вДЦ 483G, 51 pp.
- Moller P., Alexanderson H., Funder S., Hjort Ch. The Taimyr Peninsula and the Severnaya Zemlya archipelago, Arctic Russia: a synthesis of glacial history and palaeo-environmental change during the Last Glacial cycle (MIS 5ee2) // Quaternery Science Reviews. 2015. Vol.107.No 1. P.p. 149-181/
- –≠–ї–µ–Ї—В—А–Њ–љ–љ—Л–є —А–µ—Б—Г—А—Б: https://en.wikipedia.org/wiki/Mya_truncata#/media/File:Mya-truncata_in-situ_hg.jpg
{kind=link}