
Разрезы К18-15 и К18-16, пойменные образования правого берега долины Колесдален, Шпицберген
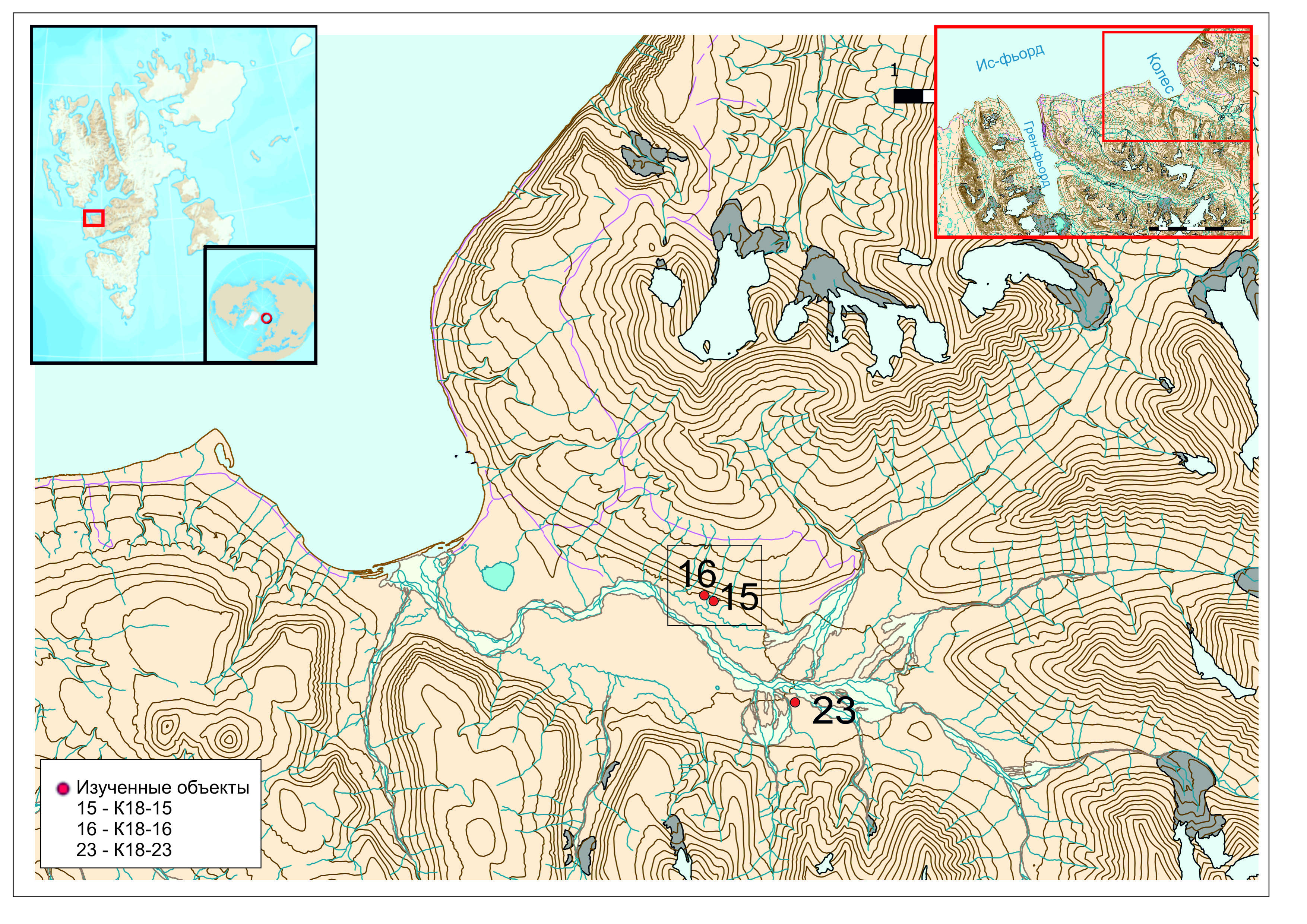
Рис. Местоположение объекта исследования
Для выявления особенностей эволюции долины Колесдален в голоцене были использованы материалы, полученные в ходе полевых работ в июле-августе 2018 года. В ходе исследований были проведены радиоуглеродный и микропалеонтологические анализы проб разрезов речных (К18-15 и К18-16) и болотных образований (К18-23). Разрезы К18-15 и К18-16 расположены в среднем течении долины р. Колес на останцах первой надпойменной террасы на правом берегу р. Колес (координаты: 78°5'41,615"N, 15°12'34,308"E - К18-15; 78°5'43,351"N, 15°12'14,865"E - К18-16;), на высоте 22 м н.у.м. (по GPS).
В разрезе К18-15 вскрыты следующие отложения (описание сверху вниз) (Рис.):
0-70 см – несортированный алевритовый материал, оборганенный с большим количеством гравия и включениями галек (преобразованный склоновыми процессами);
70 – 120 см – толща серых алевритов с тонкими прослоями – линзами хорошо разложившейся органики и пятнами ожелезнения:
105см - прослой растительный остатков, слаборазложившихся ( мощностью до 2 см)
110 см – проявление включений растительных остатков в алевритах с гравийным материалом (линзы)
В подошве пачки (вкрытых образований) – мерзлые грунты, алевриты со следами ожелезнения.

Рис. Разрез К18 – 15, литологическая колонка
В точке К18-16 поверхность останца также оголена, материал – алевриты, галька разной степени окатанности, гравий (Рис.).
В расчистке вскрыты следующие отложения (сверху вниз):
0 – 20 см алевриты бежевого цвета с охристыми пятнами, мелкоплитчатой, чешуйчатой структуры;
Около 10 см – переход к непреобразованной структуре.
18 - 22см - слабонаклонный прослой торфа коричневого цвета, хорошо разложившегося с небольшой примесью минерального компонента,
21 - 23 см – тонкий прослой гравия в торфе (мощностью до 1.5 см);
21 – 50 см - пачка неясно переслаивающихся (близко к субгоризонтальному) торфов и алевритов с прослоями м/з песка (на 30 см), включениями растительных остатков плохо разложившихся (35-40 см);
50 – 70 см торф с линзами темно-серого алеврита, чешуйчатая структура, ниже по расчистке наблюдается увеличение минерального компонента в отложениях, на глубине 95 – 100 см переход к алевритам с линзами растительных остатков торфа. Граница многолетнемерзлых пород.

Рис. Разрез К18-16, литологическая колонка
По результатам радиоуглеродного датирования было выявлено, что изученные отложения накапливались с ~ 6000 до 2900 л.н.
Методом спорово-пыльцевого анализа было изучено 10 проб из разреза К18-15 мощностью 55 см; и 12 проб из разреза К18-16, мощностью 75 см.
По результатам палинологических исследований построены две диаграммы. Процентное содержание каждого таксона рассчитано от общей суммы пыльцы наземных растений. Количество дочетвертичных микрофоссилий (Prequaternary) вычислено от общей суммы четвертичной пыльцы и спор.
В полученных спорово-пыльцевых спектрах присутствует пыльца как местных растений (Betula sect. Nanae, Salix, Cyperaceae, Poaceae, Caryophyllaceae, Ranunculaceae, Polemonium sp., Rumex sp. и др.), так и дальнезаносная (Pinus). В группе трав было определено всего 10 типов пыльцы для К18-15 и 11 типов для К18-16. Среди выделенных трав постоянно доминировали только 2 таксона – Cyperaceae и Poaceae.
По характерным изменениям в составе спорово-пыльцевых спектров и соотношению основных доминантов на диаграмме разреза К18 – 15 выделено 5 пыльцевых зон (снизу вверх) (Рис.).
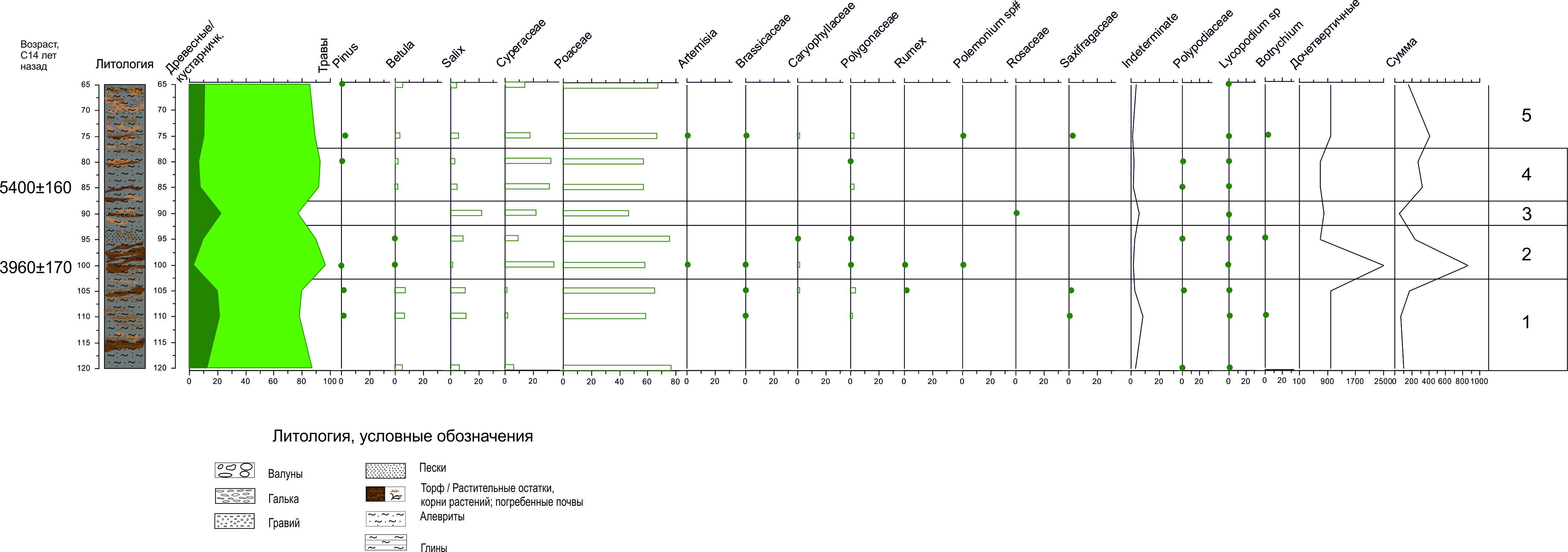
Рис. Спорово-пыльцевая диаграмма для разреза речных отложений К18-15, геохронологические данные приведены в радиоуглеродных некалиброванных л.н.
Палинозона 1 (120–102.5 см) представлена спорово-пыльцевым спектрами трех образцов (120 см, 110 см, 105 см). На фоне господства пыльцы трав (78 – 87%), содержание пыльцы Betula sect. Nanae составляет около 7 -8%, пыльцы Salix – 7 -12%. В группе трав доминирует пыльца Poaceae – 60 – 77%. Также встречается пыльца Cyperaceae (2-7%), Caryophyllaceae (4%), Brassicaceae, Rumex и др. Споры представлены в незначительном количестве такими таксонами как Huperzia sp., Sphagnum, Polypodiaceae.
Палинозона 2 (102.5 -92.5 см) представлена спектрами двух образцов (100 см, 95 см). Значительно увеличивается роль пыльцы трав (до 90 - 97%). При сохранении превосходства Poaceae 59 – 76%, содержание пыльцы Cyperaceae достигает 35%. Зафиксировано снижение концентрации пыльцы древесных. Полностью исчезает пыльца Betula sect. Nanae, количество пыльцы Salix сокращается до 2-10%. На глубине 100 см отмечено наибольшее содержание ископаемых четвертичных микрофоссилий для данного разреза.
Палинозона 3 представлена спорово-пыльцевым спектром одного образца, отобранного на глубине 90 см. Насыщенность препаратов пыльцой низкая (53 пыльцевых зерна). Значительно содержание дочетвертичных микрофоссилий. В спорово-пыльцевом спектре господствуют пыльца трав (77%), среди которой доминируют Poaceae (47%) и Cyperaceae (23%). Количество пыльцы Salix составляет 23%.
Палинозона 4 представлена спорово-пыльцевыми спектрами 2 образцов, отобранных на глубинах 80 и 85 см, где также доминирует пыльца трав (92-93%). Содержание четвертичных микрофоссилий значительно выше, чем в предыдущей зоне. Преобладает пыльца Poaceae (57-58%), Cyperaceae (32-34%). Содержание пыльцы древесных таксонов незначительно - Betula sect. Nanae 2-3%, Salix 4-6%.
Палинозона 5 также представлена спорово-пыльцевыми спектрами двух образцов (75 см, 65 см). Снижается общее количество пыльцы трав до 88-89%. По-прежнему доминирует пыльца Poaceae (67 - 70%), значительна роль Cyperaceae (15-18 %). Содержание пыльцы древесных таксонов также незначительно - Betula sect. Nanae 4 – 6 %, Salix 4-6%.
Для проб из разреза К18-16 характерна большая (по сравнению с К18-15) концентрация микрофоссилий (до 2700), их сохранность значительно лучше, количество дочетвертичных микрофоссилий сравнительно невелико. При этом, встречаются пробы с единичными зернами.
На спорово-пыльцевой диаграмме разреза К18-16 было выделено 6 палинозон (снизу вверх) (Рис.):
Палинозона 1 представлена спорово-пыльцевыми спектрами трех образцов, отобранных на глубинах 95, 80 и 75 см, и характеризуется небольшим содержанием четвертичных микрофоссилий (или практически полним их отсутствием), абсолютным преобладанием пыльцы трав, среди которых доминирует Poaceae (65-75%), среди древесных – значительна роль Betula sect. Nanae (6 -16%).
Палинозона 2 выделена по спорово-пыльцевому спектру одного образца (70 см). На фоне общего доминирования пыльцы трав, представленных в основном Poaceae (52%) и Cyperaceae (11%), значительную роль играет пыльца древесных и кустарничковых (33%), представленных Betula sect. Nanae (23 %) и Salix (10%).
Особенностью палинозоны 3 (около 5000-6000 л.н.) является высокая степень насыщенности препаратов микрофоссилиями (от 800 до 2700 зерен) с низким содержанием дочетвертичных спор и пыльцы,. На фоне господства пыльцы Cyperaceae (50 - 74%) и значительной роли пыльцы Salix (до 27%) отмечаются некоторые различия в составе спорово-пыльцевых спектров, позволивших выделить три подзоны: 3а - (65 см)- выявлено максимальное содержание пыльцы Salix (27%) и присутствием пыльца Betula sect. Nanae (2%); подзона 3b характеризуется максимальной концентрацией микрофоссилий и представлена спектрами двух образцов (60 и 55 см), в которых снижается количество пыльцы Salix до 10%, и почти полностью отсутствует пыльца Betula sect. Nanae (единичные находки). В подзоне 3с (50см) зафиксировано общее снижение концентрации микрофоссилий. При этом, количество пыльцы древесных и кустарничковых выше: Betula sect. Nanae достигает 4%, а Salix – 14%.
Палинозона 4 выделена по спорово-пыльцевому спектру одного образца, отобранного на глубине 40 см, для которого характерно абсолютное преобладание пыльцы трав (96%), среди которой доминирует Poaceae (66%), присутствует значительное количество пыльцы Cyperaceae (27%). Роль древесных таксонов невелика, присутствуют Salix (2%) и Betula sect. Nanae (2%).
Палинозона 5 (около 4000 л.н.) представлена спорово-пыльцевыми спектрами двух образцов (25 и 35 см). Насыщенность препаратов микрофоссилиями низкая. В спорово-пыльцевых спектрах доминирует пыльца трав (72-80%), при этом содержание пыльцы Poaceae колеблется от 25% до 68%, а Cyperaceae сокращается вверх по разрезу от 33% до 2%. При этом значительна роль пыльцы Betula sect. Nanae (17 – 21%), присутствует пыльца Brassicaceae, Polemonium sp., Polygonaceae, Caryophyllaceae.
Палинозона 6 (около 3000 л.н) выделена по спорово-пыльцевому спектру одного образца (20 см), для которого характерна высокая насыщенность четвертичными спорами и пыльцой, с абсолютным преобладанием пыльцы Cyperaceae (59%), значительным содержанием пыльцы Poaceae (39%). Содержание пыльцы древесных и кустарничковых не значительно и не превышает 1%.
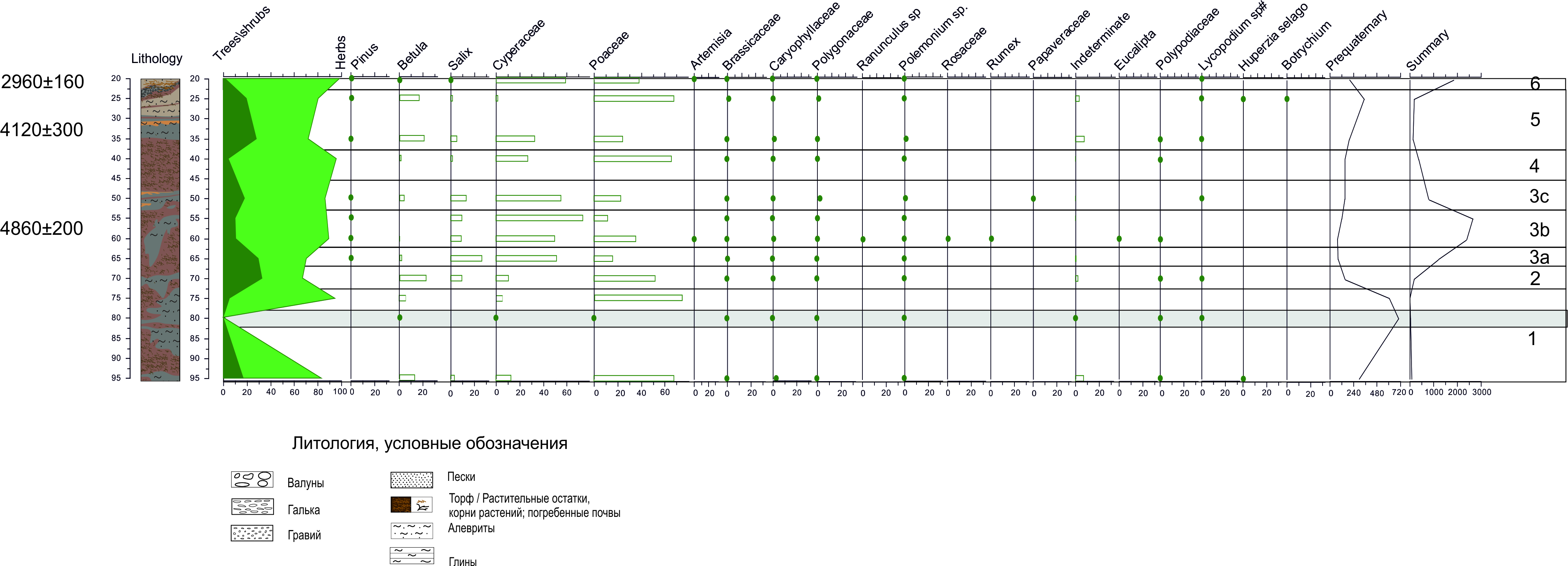
Рис. Спорово-пыльцевая диаграмма для разреза речных отложений К18-16, геохронологические данные приведены в радиоуглеродных некалиброванных л.н.
В разрезе установлены численно богатые бентосные пресноводные диатомовые комплексы, состоящие из 30 видов и немногочисленные цисты золотистых водорослей.
Поскольку по всей мощности разреза преобладают пресноводные бентосные виды, для наблюдения за динамикой диатомовых комплексов была выбрана характеристика реакции среды (рН). В диаграмме отражено соотношение ацидофильных, алкалофильных и индифферентных к реакции рН видов (Рис.).
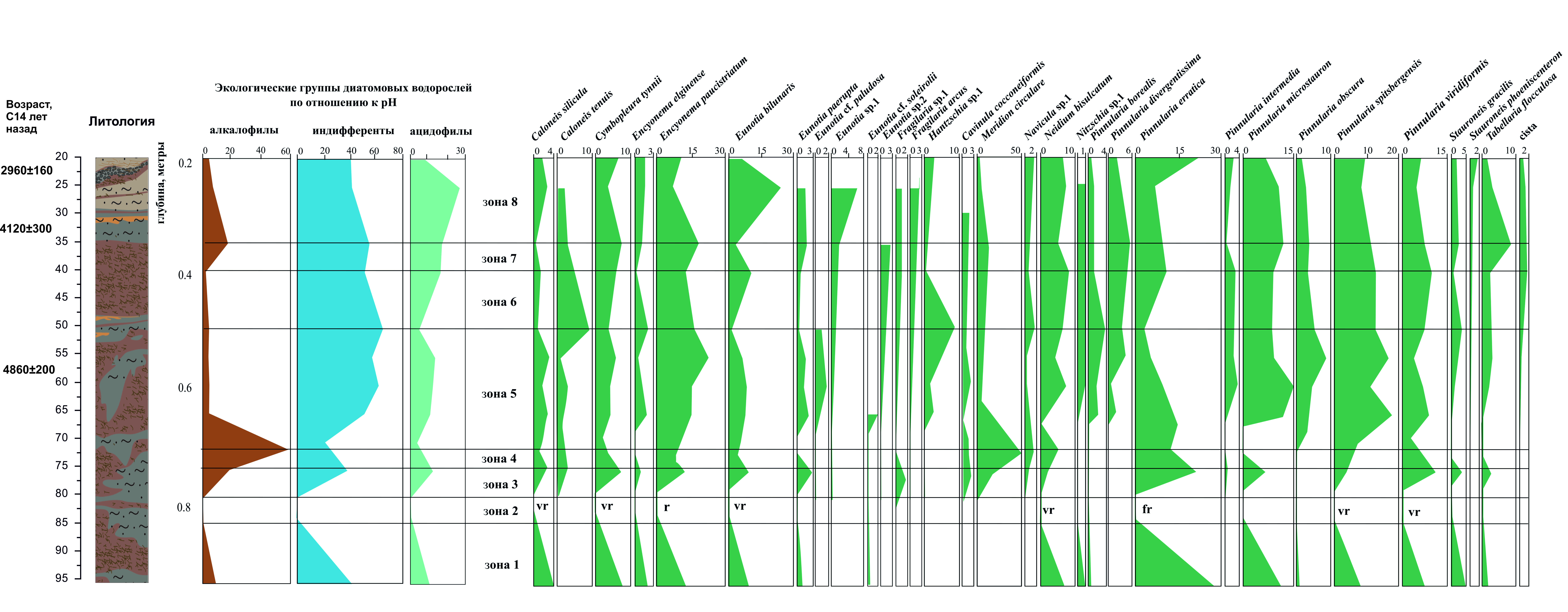
Рис. Диаграмма распределения видов диатомовых водорослей по глубине разреза К18-16, геохронологические данные приведены в радиоуглеродных некалиброванных л.н.
Всего выделено 8 зон (Рис.):
Зона 1 установлена на глубине 80-95 см, где преобладают бентосные пресноводные индифферентные по отношению к pH виды. Доминирует донный олиготрофный рН-индифферентный вид Pinnularia erratica. В том числе присутствуют пресноводные донные виды: алкалофильный Caloneis silicula, индифферентные виды Pinnularia microstauron и Pinnularia viridiformis. Присутствуют арктические виды Cymbopleura tynnii и Pinnularia spitsbergensis; а также, олиготрофные виды Encyonema paucistriatum и Neidium bisulcatum. Ацидофильные виды диатомовых водорослей составляют 12%, к ним относятся виды Eunotia bilunaris и Tabellaria flocculosa.
Формирование этого горизонта отложений происходило в условиях холодного пресного неглубокого водоёма с нейтральной реакцией среды, в который, вероятно, поступали болотные воды.
Зона 2 установлена на глубине 80 см и представлена численно бедным комплексом с преобладанием диатомового вида Pinnularia erratica, также, в виде отдельных створок встречены виды Encyonema paucistriatum, Caloneis silicula, Cymbopleura tynnii, Eunotia bilunaris, Neidium bisulcatum, Pinnularia viridiformis и Pinnularia spitsbergensis.
Формирование этого горизонта отложений происходило в неблагоприятных условиях холодного пресного неглубокого водоёма с нейтральной реакцией среды.
Зона 3 соответствует глубине 75-80 см и отличается увеличением количества всех преобладающих видов диатомовых водорослей – доминантного вида Pinnularia erratica, преобладающих видов Encyonema paucistriatum, Caloneis silicula, Cymbopleura tynnii, Eunotia bilunaris, Neidium bisulcatum, Pinnularia viridiformis, Pinnularia microstauron и Pinnularia spitsbergensis. Начиная с этой глубины к преобладающим видам добавляются следующие: ацидофильный арктический вид Eunotia praerupta, алкалофильный вид Cavinula cocconeiformis, пресноводный реофильный вид Meridion circulare и Fragilaria sp.
Формирование этого горизонта отложений происходило в переходных условиях пресного неглубокого водоёма, в котором начинает изменяться реакция среды.
Зона 4 соответствует глубине 70-75 см и характеризуется высоким содержанием алкалофильных видов (до 40%) и доминированием пресноводного алкалофильного реофильного вида Meridion circulare. Одновременно с этим прослеживается снижение количества пресноводных видов рода Pinnularia. Остальные виды присутствуют, но их количество меньше по сравнению с зоной 3.
Формирование этого горизонта отложений происходило в условиях пресного неглубокого водоёма с реакцией среды от нейтральной до слабощелочной. Возможно, с попаданием проточных вод, что может указывать на увеличение стока.
Зона 5 зафиксирована на глубине 50-70 см, характеризуется снижением количества алкалофильных видов и ростом количества индифферентов. Преобладают виды рода Pinnularia: донный вид P. microstauron, болотный вид P. obscura и арктический вид P. spitsbergensis (до 17%) на фоне снижения количества вида P. erratica. Доминирует олиготрофный вид-обрастатель Encyonema paucistriatum. Ацидофильные виды составляют до 15% - виды рода Eunotia, в том числе арктического вида Eunotia praerupta, и Tabellaria flocculosa. Обнаруживается вид Hantzschia sp.
Формирование этого горизонта отложений происходило в условиях холодного пресного неглубокого водоёма с нейтральной реакцией среды. Наличие до 15% ацидофильных видов указывает на участие болотных вод, а большое количество створок арктических видов (20% и более) - на холодные условия среды.
Зона 6 соответствует глубине 40-50 см, характеризуется преобладанием индифферентных видов с тенденцией к снижению их количества по разрезу, ростом количества ацидофильных видов (до 18% на глубине 40 см), а также почти полным отсутствием алкалофильных видов (<5%). Преобладают следующие виды: индифферентный пресноводный вид Caloneis tenuis, олиготрофный вид-обрастатель Encyonema paucistriatum, вид Neidium bisulcatum и виды рода Pinnularia (P. erratica, P. microstauron, P. spitsbergensis, P. viridiformis, P. obscura). Количество створок арктических видов (Pinnularia spitsbergensis, Eunotia praerupta, Cymbopleura tynnii) достигает 18-20%.
Формирование этого горизонта отложений происходило в условиях холодного пресного неглубокого водоёма с нейтральной реакцией среды и тенденцией к заболачиванию.
Зона 7 соответствует глубине 30-40 см. На этих глубинах зафиксировано кратковременное повышение количества алкалофильных видов до 20% при наличии 20-25% ацидофильных видов. Помимо преобладания пресноводных видов рода Pinnularia, увеличивают концентрацию следующие виды: арктический олиготрофный вид, часто встречающийся на мхах - Cymbopleura tynnii, пресноводный ацидофильный вид Tabellaria flocculosa, олиготрофный вид-обрастатель Encyonema paucistriatum и ацидофильный олиготрофный Pinnularia divergentissima.
Формирование этого горизонта отложений происходило в условиях холодного пресного мелкого водоёма со сложной реакцией среды.
Зона 8 зафиксирована на глубине 20-30 см и характеризуется повышением количества ацидофильных видов до 30%. Преобладают следующие диатомовые виды: пресноводный ацидофильный вид-эвритоп Eunotia bilunaris, виды рода Pinnularia - P. erratica, P. microstauron, P. spitsbergensis, пресноводный олиготрофный вид Neidium bisulcatum. олиготрофный вид-обрастатель Encyonema paucistriatum.
Формирование этого горизонта отложений происходило в условиях пресного мелкого водоёма с нейтральной до слабокислой реакцией среды и процессами заболачивания.
По полученным данным, для среднего течения р. Колес можно выделить следующие особенности эволюции растительности, отображенные в последовательности сменяющих друг друга этапов:
1 этап - формирование отложений в нижней части разреза (120 – 90 см) – отражает развитие кустарничково-травяных тундр. Участие кустарничковых ив и берез значительно, среди трав лидируют злаковые и осоковые. Преобладание ивки полярной среди древесных и кустарничковых свидетельствует о значительной снежности (накопление достаточного снежного покрова в холодный сезон). Вероятно, период формирования толщи характеризовался относительно теплыми и влажными условиями, с формированием устойчивого снежного покрова.
Низкая концентрация четвертичных микрофоссилий в отложениях, соответствующих этапу 2 (90 – 75 см) до ~ 6000 - 5500 л.н., может свидетельствовать о неблагоприятных климатических условиях. При этом в препаратах по-прежнему преобладают злаковые. Увеличение концентраций пыльцы поляной ивки и уменьшение березы также отражают похолодание. Вероятнее всего, в этот период сформировались ивово-травяные сообщества в относительно прохладном и более влажном климате. По данным диатомового анализа в этот этап формирование отложений происходило в условиях холодного пресного неглубокого водоёма с нейтральной реакцией среды, в который, вероятно, поступали болотные воды.
На следующем этапе (75 – 70 см) ~ 5500 – 5000 л.н. при общем сохранении в растительности основных преобладающих таксонов (злаковые), условия осадконакопления были значительно благоприятнее, о чем говорит рост концентрации четвертичных микрофоссилий и увеличение роли пыльцы карликовой березки. Зафиксировано увеличение содержания растительных остатков в отложениях. Господствовала кустарничково-травяная тундра. Изменение соотношения видов диатомовых водорослей с разной реакцией рН свидетельствует о начале процессов заболачивания. Спектр диатомовых водорослей, изученных в образце на глубине 70 см фиксирует резкое увеличение рН среды и присутствие проточных вод. Это может указывать на увеличение стока вследствие кратковременного потепления в период накопления отложений.
Увеличение концентрации микрофоссилий, количества растительных остатков в отложениях на этапе 4 (70 – 45см) ~ 4500 - 5000 л.н., а также постепенная смена доминирующих таксонов свидетельствует о перестройке природной среды. На данном этапе условия для сохранения четвертичных спор и пыльцы были наиболее благоприятными, формировались относительно мощные торфяные горизонты. В растительном покрове преобладают осоковые, роль злаковых значительно меньше. Среди древесных и кустарничковых значительна роль ивки, что свидетельствует о высокой степени увлажнения. Данные о диатомовых комплексах подтверждают полученные выводы. Отложения формировались в условиях холодного пресного неглубокого водоёма с нейтральной реакцией среды и тенденцией к заболачиванию.
Обстановка осадконакопления на этапе 5 (~4500 – 3000 л.н.) была менее благоприятной для сохранения пыльцы и спор. Вероятно, это связано с изменением динамики среды. Тем не менее, можно сделать вывод, растительный покров на этом этапе представлял собой кустарничковую (березовую) злаково-осоковую и осоково – злаковую тундру с присутствием ивки.
На этапе 6 (~ 3000 л.н.), развивалась злаково-осоковая тундра. Количество кустарничковых значительно меньше, вероятно, наличие их пыльцы является фоновым показателем.
1. Сурова Т.Г., Троицкий Л.С., Пуннинг Я.-М.К. Об истории оледенения Шпицбергена в голоцене по данным палеоботанических исследований // Материалы гляциологических исследований, 1982. № 42. С. 100-106.
Электронные ресурсы:
2. Норвежский полярный институт - Norwegian Polar Institute [Электронный ресурс https://geokart.npolar.no/Html5Viewer/index.html?viewer=Svalbardkartet, свободный, (дата обращения: 30.05.20).
3. Карты Шпицбергена - Map of Svalbard [Электронный ресурс http://toposvalbard.npolar.no/, свободный, (дата обращения: 03.06.20).
4. Программа С2 для построения спорово-пыльцевых диаграмм [Электронный ресурс]. – Режим доступа: www.staff.ncl.ac.uk/stephen.juggins/software/C2Home.htm/, свободный, (дата обращения: 17.05.20).